UNIDAD I INTRODUCCIÓN A LA FISIOLOGÍA: LA CÉLULA Y LA FISIOLOGÍA GENERAL
Organización funcional del cuerpo humano y control del «medio interno»

La fisiología es la ciencia que pretende explicar los mecanismos físicos y químicos responsables del origen, desarrollo y progresión de la vida. La mayoría de las funciones fisiológicas pueden separarse en fisiología: vírica, bacteriana, celular, vegetal, de los invertebrados, de los vertebrados, de los mamíferos, humana y muchas otras subdivisiones.
Fisiología humana
La fisiología humana intenta explicar las características y mecanismos específicos del cuerpo humano que hacen que sea un ser vivo. El hecho de mantenerse vivo es el resultado de sistemas de control complejos.
Las células como unidades vivas del cuerpo

La unidad viva básica del cuerpo es la célula. Cada órgano es un agregado de muchas células diferentes que se mantienen unidas mediante estructuras de soporte intercelulares. Cada tipo de célula está especialmente adaptado para realizar una o más funciones concretas.
Por ejemplo, los eritrocitos, cuya cantidad asciende aproximadamente a 25 billones en cada ser humano, transportan el oxígeno desde los pulmones a los tejidos. Aunque los eritrocitos son las más abundantes entre todas las células corporales, hay 75 billones de células más de otros tipos que realizan funciones diferentes. El cuerpo en su conjunto contiene en torno a 100 billones de células. Aunque las múltiples células del cuerpo son muy diferentes entre sí, todas ellas tienen determinadas características básicas que son similares.

Los mecanismos químicos generales que permiten cambiar los nutrientes en energía son básicamente los mismos en todas las células y todas las células liberan los productos de sus reacciones químicas en los líquidos circundantes. Todas las células tienen la capacidad de reproducirse formando más células de su propia estirpe. Por fortuna, cuando se destruyen células de un tipo en particular, el resto de las células de este tipo genera nuevas células hasta rellenar el cupo.
Líquido extracelular: el «medio interno»
El 60% del cuerpo humano del adulto es líquido, principalmente una solución acuosa de iones y otras sustancias. Si bien casi todo este líquido queda dentro de las células y se conoce como líquido intracelular, aproximadamente una tercera parte se encuentra en los espacios exteriores a las células y se denomina líquido extracelular.
Este líquido extracelular está en movimiento constante por todo el cuerpo y se transporta rápidamente en la sangre circulante para mezclarse después entre la sangre y los líquidos tisulares por difusión a través de las paredes capilares. En el líquido extracelular están los iones y nutrientes que necesitan las células para mantenerse vivas, por este motivo, el líquido extracelular también se denomina medio interno del organismo.
Las células son capaces de vivir y realizar sus funciones especiales, siempre que este medio interno disponga de las concentraciones adecuadas de oxígeno, glucosa, distintos iones, aminoácidos, sustancias grasas y otros componentes.
Diferencias entre los líquidos extracelular e intracelular
Líquido extracelular: contiene grandes cantidades de iones sodio, cloruro y bicarbonato más nutrientes para las células, como oxígeno, glucosa, ácidos grasos y aminoácidos. También contiene dióxido de carbono, que se transporta desde las células a los pulmones para ser excretado junto con otros residuos celulares que se transportan a los riñones para su excreción.
Líquido intracelular: contiene grandes cantidades de iones potasio, magnesio y fosfato.
Los mecanismos especiales de transporte de iones a través de la membrana celular mantienen las diferencias en la concentración de iones entre los líquidos extracelular e intracelular.
Homeostasis: mantenimiento de un medio interno casi constante
En 1929, el fisiólogo estadounidense walter cannon (1871-1945) acuñó el término homeostasis para referirse al mantenimiento de unas condiciones casi constantes del medio interno. Esencialmente todos los órganos y tejidos del organismo realizan funciones que colaboran en el mantenimiento de estas condiciones relativamente constantes.
Los diversos iones, nutrientes, productos de desecho y otros componentes del organismo están regulados normalmente dentro de un intervalo de valores, no poseen valores fijos. Existen poderosos sistemas de control para mantener las concentraciones de sodio e hidrógeno, así como la mayoría de los demás iones, nutrientes y sustancias del organismo, en niveles que permitan que las células, los tejidos y los órganos lleven a cabo sus funciones normales, pese a grandes variaciones ambientales y a las dificultades derivadas de lesiones y enfermedades.
Las funciones normales del organismo exigen acciones integradas de células, tejidos, órganos y los múltiples sistemas de control nervioso, hormonales y locales que contribuyen conjuntamente a la homeostasis y a la buena salud.
La enfermedad se considera un estado de ruptura de la homeostasis. Sin embargo, incluso en presencia de enfermedades, los mecanismos homeostáticos siguen activos y mantienen las funciones vitales a través de múltiples compensaciones.
Las compensaciones homeostáticas que se producen en el organismo después de una lesión, una enfermedad o de cambios ambientales importantes pueden verse como un «compromiso» necesario para mantener las funciones vitales si bien, a largo plazo, pueden contribuir a inducir anomalías adicionales en el organismo. La disciplina de la fisiopatología pretende explicar cómo se alteran los diversos procesos fisiológicos durante las enfermedades y las lesiones.
Transporte en el líquido extracelular y sistema de mezcla: el aparato circulatorio
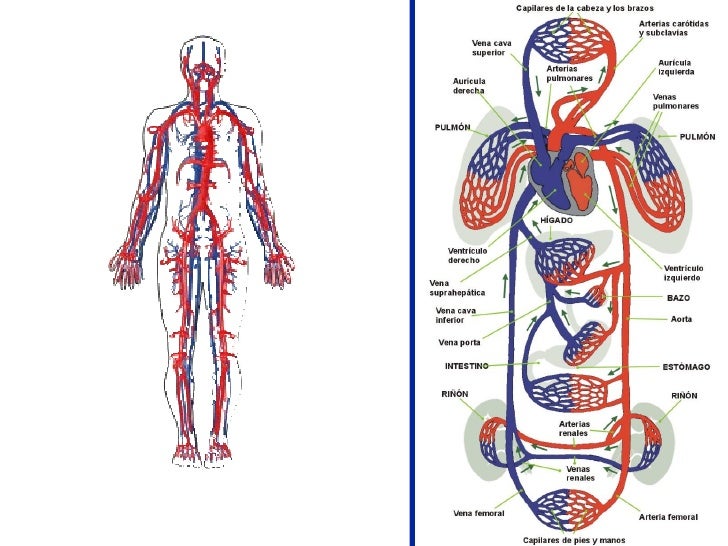
El líquido extracelular circula por el organismo en dos etapas:
Primera etapa: consiste en el movimiento de la sangre por el cuerpo dentro de los vasos sanguíneos.
Segunda etapa: es el movimiento del líquido entre los capilares sanguíneos y los espacios intercelulares entre las células tisulares.
A medida que la sangre atraviesa los capilares sanguíneos se produce también un intercambio continuo de líquido extracelular entre la porción del plasma de la sangre y el líquido intersticial que rellena los espacios intercelulares. Las paredes de los capilares son permeables a la mayoría de las moléculas del plasma sanguíneo, con la excepción de las proteínas plasmáticas, que son demasiado grandes para pasar con facilidad a través de los capilares. Grandes cantidades de líquido y sus componentes disueltos difunden yendo y viniendo entre la sangre y los espacios tisulares. Este proceso de difusión se debe al movimiento cinético de las moléculas en el plasma y en el líquido intersticial, es decir, el líquido y las moléculas disueltas están en movimiento continuo y van dando tumbos en todas las direcciones dentro del plasma y el líquido en los espacios intercelulares, además de atravesar los poros capilares.
Origen de los nutrientes en el líquido extracelular
Aparato respiratorio
Cada vez que la sangre atraviesa el organismo también fluye por los pulmones y capta el oxígeno a través de los alvéolos, adquiriendo el oxígeno que necesitan las células. La membrana que separa los alvéolos y la luz de los capilares pulmonares, la membrana alveolar, tiene un grosor de tan solo 0,4 a 2 μm y el oxígeno difunde rápidamente por el movimiento molecular a través de esta membrana para entrar en la sangre.
Aparato digestivo
Una gran porción de la sangre que bombea el corazón también atraviesa las paredes del aparato digestivo, donde se absorben los distintos nutrientes, incluidos los hidratos de carbono, los ácidos grasos y los aminoácidos, desde el alimento ingerido hacia el líquido extracelular de la sangre.
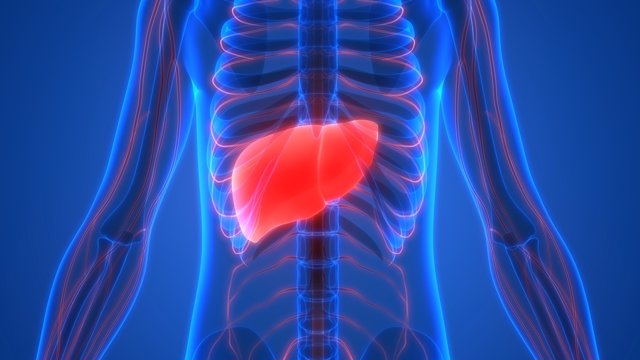
Hígado y otros órganos que realizan principalmente funciones metabólicas
No todas las sustancias absorbidas del aparato digestivo pueden usarse tal como las células las absorben y el hígado es el encargado de cambiar la composición química de muchas de ellas, para convertirlas en formas más utilizables, mientras que otros tejidos corporales, los adipocitos, la mucosa digestiva, los riñones y las glándulas endocrinas, modifican o almacenan las sustancias absorbidas hasta que son necesitadas. El hígado elimina también ciertos residuos producidos en el cuerpo y las sustancias tóxicas que se ingieren.
Aparato locomotor
Si no fuera por los músculos, el organismo no podría desplazarse para obtener los alimentos que se necesitan para la nutrición. El aparato locomotor también permite la movilidad como protección frente al entorno, sin la cual todo el organismo, incluidos sus mecanismos homeostáticos, sería destruido.

Eliminación de los productos finales metabólicos
Eliminación del dióxido de carbono en los pulmones
Al mismo tiempo que la sangre capta el oxígeno en los pulmones, se libera el dióxido de carbono desde la sangre hacia los alvéolos y el movimiento respiratorio de aire que entra y sale de los pulmones transporta el dióxido de carbono hacia la atmósfera. El dióxido de carbono es el más abundante de todos los productos del metabolismo.
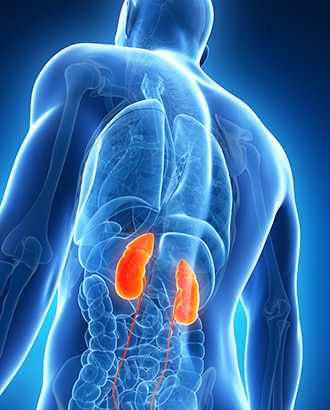
Riñones
Con el paso de la sangre a través de los riñones se eliminan del plasma la mayoría de las sustancias que las células ya no necesitan, como son los distintos productos finales del metabolismo celular, como: la urea y el ácido úrico y el exceso de iones y agua de los alimentos, que podrían acumularse en el líquido extracelular. La mayoría de las demás sustancias que el organismo no necesita (en especial los productos de desecho metabólicos) se reabsorben mal y atraviesan los túbulos renales hacia la orina. Los riñones realizan su función filtrando primero una gran cantidad de plasma a través de los capilares de los glomérulos hacia los túbulos y reabsorbiendo hacia la sangre las sustancias que necesita el organismo, como: glucosa, aminoácidos, cantidades apropiadas de agua y muchos de los iones.
Aparato digestivo
El material no digerido que entra en el aparato digestivo y algunos productos residuales del metabolismo se eliminan en las heces.
Hígado
Entre sus funciones se encuentra la detoxificación o eliminación de numerosos fármacos y productos químicos que se ingieren. Secreta muchos de estos residuos en la bilis para su eliminación ulterior en las heces.
Regulación de las funciones corporales
Sistema nervioso
Está compuesto por tres partes principales:
- La porción de aferencia sensitiva. Los receptores sensitivos detectan el estado del cuerpo o de su entorno (piel, ojos, oidos, etc).
- Sistema nervioso central (o la porción integradora). Está formado por el cerebro y la médula espinal. El cerebro almacena información, genera los pensamientos, crea la ambición y determina las reacciones que debe manifestar el cuerpo en respuesta a las sensaciones para, a continuación, transmitir las señales apropiadas a través de la porción motora eferente del sistema nervioso.

Un segmento importante del sistema nervioso es el sistema nervioso autónomo o neurovegetativo, que funciona a escala subconsciente y controla muchas de las funciones de los órganos internos, como la función de bomba del corazón, los movimientos del aparato digestivo y la secreción en muchas de las glándulas corporales.
3.La porción eferente motora. Llevar a cabo los deseos del sujeto
Sistemas hormonales
Dentro del organismo se encuentran ocho glándulas endocrinas mayores y varios órganos y tejidos que segregan productos químicos denominados hormonas. Las hormonas se transportan en el líquido extracelular a otras partes del cuerpo para regular las funciones celulares, proporcionan un sistema de regulación que complementa al sistema nervioso. Ejemplo:
Hormona tiroidea: aumenta la velocidad de la mayoría de las reacciones químicas de todas las células, con lo que se facilita el ritmo de la actividad corporal.
Insulina: controla el metabolismo de la glucosa.
Hormonas corticosuprarrenales: controlan los iones sodio y potasio y el metabolismo proteico.

Hormona paratiroidea: controla el calcio y el fosfato en el hueso.
El sistema nervioso regula numerosas actividades musculares y secretoras del organismo, mientras que el sistema hormonal regula muchas de las funciones metabólicas. Normalmente, los sistemas nerviosos y hormonales trabajan de forma coordinada para controlar esencialmente todos los sistemas orgánicos del cuerpo.
Protección del cuerpo
Sistema inmunitario
El sistema inmunitario está formado por los glóbulos blancos, células tisulares derivadas de los glóbulos blancos, el timo, los nódulos linfáticos y los vasos linfáticos que protegen el cuerpo de patógenos como bacterias, virus, parásitos y hongos.
El sistema inmunitario proporciona un mecanismo para que el cuerpo:
1) diferencie sus propias células de las células y sustancias extrañas.
2) destruya al invasor por fagocitosis o mediante la producción de linfocitos sensibilizados o proteínas especializadas (p. Ej., anticuerpos) que destruyen o neutralizan al invasor.

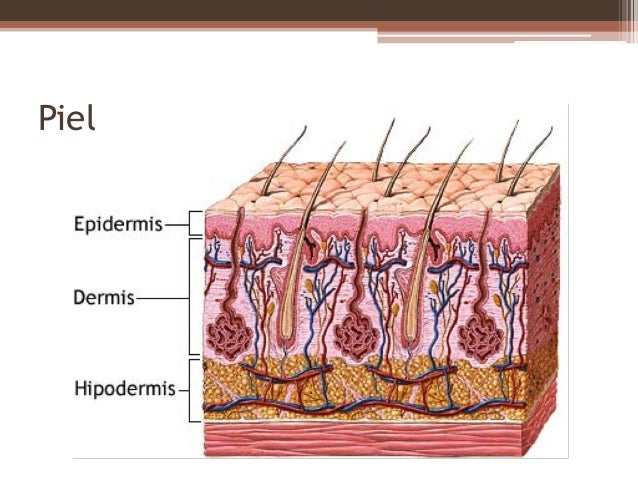
Sistema tegumentario
La piel y sus diversos anejos, como el pelo, las uñas, las glándulas y otras estructuras, cubren, amortiguan y protegen los tejidos profundos y los órganos del cuerpo y, en general, definen una frontera entre el medio corporal interno y el mundo exterior. Es importante también para la regulación de la temperatura y la excreción de los residuos y proporciona una interfaz sensorial entre el cuerpo y el medio exterior. La piel suele comprender entre aproximadamente el 12 y el 15% del peso corporal.
Reproducción
A veces no se considera que la reproducción sea una función homeostática, aunque ayuda a mantener la homeostasis generando nuevos seres que ocuparán el lugar de aquellos que mueren.
Sistemas de control del organismo
El cuerpo humano contiene miles de sistemas de control. Algunos de los más intrincados de estos sistemas son los de control genético que actúan en todas las células para mantener el control de las funciones intracelulares y extracelulares. Hay muchos otros sistemas de control que actúan dentro de los órganos para controlar las funciones de sus componentes, otros actúan a través de todo el organismo para controlar las interrelaciones entre los órganos.
Ejemplos de mecanismos de control
Regulación de las concentraciones de oxígeno y dióxido de carbono en el líquido extracelular
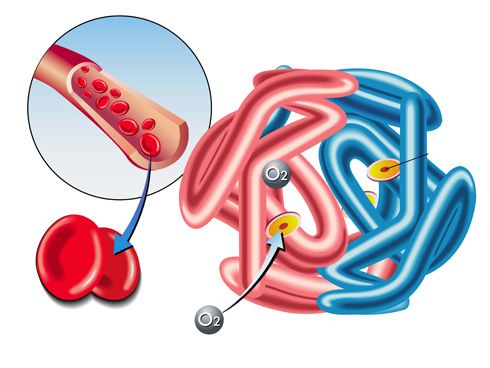
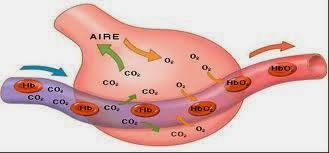
Como el oxígeno es una de las principales sustancias que requieren las reacciones químicas de las células, el organismo tiene un mecanismo de control especial para mantener una concentración casi exacta y constante de oxígeno en el líquido extracelular. Este mecanismo depende principalmente de las características químicas de la hemoglobina, que está presente en todos los eritrocitos. La hemoglobina se combina con el oxígeno a medida que la sangre atraviesa los pulmones. Posteriormente, cuando la sangre atraviesa los capilares tisulares, su propia afinidad química por el oxígeno permite que no lo libere en los tejidos si ya hay demasiado. Sin embargo, si la concentración de oxígeno en el líquido tisular es demasiado baja, se libera oxígeno suficiente para restablecer una concentración adecuada. La regulación de la concentración de oxígeno en los tejidos se conoce como función amortiguadora de oxígeno de la hemoglobina. El dióxido de carbono es el principal producto final de las reacciones oxidativas de las células; si todo el dióxido de carbono que se forma en ellas se acumulara en los líquidos tisulares, todas las reacciones que aportan oxígeno a la célula cesarían. Por fortuna, una concentración mayor de lo normal de dióxido de carbono en la sangre excita el centro respiratorio, haciendo que la persona tenga una respiración rápida y profunda. Esta aumenta la espiración de dióxido de carbono y, por tanto, elimina el exceso de dióxido de carbono de la sangre y los líquidos tisulares. Este proceso continúa hasta que la concentración vuelve a la normalidad.
Regulación de la presión arterial
Hay varios sistemas que contribuyen a la regulación de la presión arterial. Uno de ellos, el sistema de barorreceptores es un mecanismo de control de acción rápida.
En las paredes de la zona en que se bifurcan las arterias carótidas en el cuello, y también en el cayado aórtico en el tórax, se encuentran muchos receptores nerviosos denominados barorreceptores que se estimulan cuando se estira la pared arterial. Cuando la presión arterial es demasiado elevada los barorreceptores envían descargas de impulsos nerviosos al bulbo raquídeo cerebral, que es donde estos impulsos inhiben el centro vasomotor y, a su vez, disminuyen el número de impulsos transmitidos desde el centro vasomotor a través del sistema nervioso simpático hacia el corazón y los vasos sanguíneos. La ausencia de estos impulsos hace que disminuya la actividad de bomba en el corazón y también produce una dilatación de los vasos sanguíneos periféricos, lo que permite aumentar el flujo de sangre a través de ellos. Ambos efectos hacen que la presión arterial disminuya y tienda a recuperar sus valores normales. Por el contrario, el descenso de la presión arterial por debajo de lo normal relaja los receptores de estiramiento y hace que el centro vasomotor se vuelva más activo de lo habitual, con lo que se provoca vasoconstricción y un aumento de la acción de la bomba cardíaca. Así, el descenso en la presión arterial conlleva también una elevación hasta alcanzar la normalidad.
Valores normales y características físicas de los principales componentes del líquido extracelular
Obsérvese que el intervalo normal de cada uno de ellos es muy estrecho. Los valores fuera de estos intervalos suelen deberse a una enfermedad, una lesión u otros problemas importantes en el medio. Lo más importante es conocer los límites por encima de los cuales estas alteraciones provocan la muerte. Por ejemplo, siempre que disminuya a menos de un tercio de la normalidad de la concentración del ion potasio es probable que la persona quede paralizada debido a que los nervios ya no pueden transportar las señales. Por el contrario, cuando la concentración del ion potasio aumenta dos o más veces por encima de lo normal es probable que el músculo cardíaco esté muy deprimido. Cuando la concentración del ion calcio se reduce a la mitad de la normalidad aparecen contracciones tetánicas de los músculos de todo el cuerpo por la generación espontánea de un número excesivo de impulsos nerviosos en los nervios periféricos. Cuando la concentración de glucosa disminuye por debajo de la mitad de lo normal, se desarrolla una irritabilidad mental extrema y, en ocasiones, incluso aparecen convulsiones.
Características de los sistemas de control
Retroalimentación negativa de la mayoría de los sistemas de control

La mayoría de los sistemas de control del organismo actúan mediante una retroalimentación negativa. Al hablar de la regulación de la concentración del dióxido de carbono, la ventilación pulmonar aumenta cuando dicha concentración se eleva en el líquido extracelular. A su vez, el aumento de la ventilación pulmonar disminuye la concentración de dióxido de carbono en el líquido extracelular porque los pulmones espiran cantidades mayores de dióxido de carbono del organismo. La concentración elevada de dióxido de carbono inicia una serie de sucesos que disminuyen la concentración hacia la normalidad, lo que es una señal negativa para iniciar el estímulo. Por el contrario, una concentración de dióxido de carbono que disminuye demasiado produce una retroalimentación que tiende a aumentar la concentración. Esta respuesta también es negativa para iniciar el estímulo. Por tanto, en general, si algún factor se vuelve excesivo o deficiente, un sistema de control inicia una retroalimentación negativa que consiste en una serie de cambios que devuelven ese factor hacia un determinado valor medio, con lo que se mantiene la homeostasis.
La retroalimentación positiva a veces provoca círculos viciosos y la muerte
Si se tiene en cuenta la naturaleza de la retroalimentación positiva, resulta evidente que no consigue la estabilidad, sino la inestabilidad y, en algunos casos, puede causar la muerte. Ejemplo: el corazón de un ser humano sano bombea aproximadamente 5 l de sangre por minuto. Si una persona tiene bruscamente una hemorragia de 2 l, la cantidad de sangre del organismo disminuye hasta un nivel tan bajo que no queda sangre suficiente para que el corazón bombee eficazmente. En consecuencia, cae la presión arterial y disminuye el flujo de sangre que llega hacia el músculo cardíaco a través de los vasos coronarios. Este escenario lleva a que el corazón se debilite, el efecto de bomba pierda eficacia, disminuya aún más el flujo de sangre coronario y el corazón se debilite aún más; este ciclo se repite una y otra vez, hasta que se produce la muerte. La retroalimentación positiva se debería denominar mejor «círculo vicioso», aunque los mecanismos de control de retroalimentación negativa del organismo pueden superar los grados leves de retroalimentación positiva y no se desarrolla el círculo vicioso.
La retroalimentación positiva a veces es útil
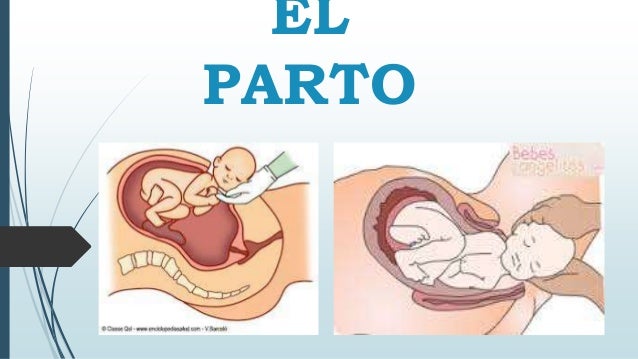
En algunos casos, el organismo usa la retroalimentación positiva a su favor. La coagulación sanguínea, el parto y la generación de señales nerviosas son ejemplos del gran valor que tiene la retroalimentación positiva.
Cuando se rompe un vaso sanguíneo y comienza a formarse un coágulo, dentro de este se activan muchas enzimas denominadas factores de coagulación. Algunas de estas enzimas actúan sobre otras enzimas inactivadas que están en la sangre inmediatamente adyacente, con lo que se consigue que coagule más sangre. Este proceso continúa hasta que el orificio del vaso se tapona y cesa la hemorragia. A veces, este mecanismo se descontrola y provoca la formación de coágulos no deseados. Este proceso es el que inicia la mayoría de los ataques cardíacos.
Cuando las contracciones uterinas son suficientemente fuertes como para que la cabeza del niño comience a empujar el cuello uterino, el estiramiento de este envía señales a través del músculo uterino que vuelven hasta el cuerpo del útero, provocando contracciones aún más potentes. Es decir, las contracciones uterinas estiran el cuello y el estiramiento del cuello provoca contracciones más potentes. El niño nace cuando este proceso adquiere la potencia suficiente; si no lo hace, las contracciones se desvanecen y transcurren algunos días hasta que vuelven a comenzar.
La estimulación de la membrana de una fibra nerviosa provoca una pequeña pérdida de iones sodio a través de los canales de sodio de la membrana nerviosa hacia el interior de la fibra. Los iones sodio que entran en la fibra cambian el potencial de membrana, lo que a su vez provoca la apertura de más canales, un cambio mayor del potencial, la apertura de más canales, y así sucesivamente. Es decir, una pequeña fuga se convierte en una explosión de sodio que entra en la fibra nerviosa creando un potencial de acción en el nervio. Este potencial de acción provoca, a su vez, una corriente eléctrica que fluye a lo largo del exterior y del interior de la fibra nerviosa e inicia nuevos potenciales de acción. Este proceso continúa una y otra vez hasta que la señal nerviosa recorre la fibra hasta su extremo. Siempre que la retroalimentación positiva es útil, la retroalimentación positiva forma parte de un proceso global de retroalimentación negativa. Además, la retroalimentación positiva que provoca las señales nerviosas permite que los nervios participen en los miles de sistemas de control de retroalimentación negativa de los nervios.
Tipos más complejos de sistemas de control: control adaptativo
Algunos movimientos del organismo son tan rápidos que no hay tiempo suficiente para que las señales nerviosas se desplacen desde la periferia del organismo hasta el cerebro y vuelvan a la periferia para controlar el movimiento, por lo que el cerebro aplica un principio que se conoce como control anterógrado, que hace que se contraigan los músculos apropiados, es decir, las señales del nervio sensible de las partes en movimiento informan al cerebro si el movimiento se está realizando correctamente. En caso contrario, el cerebro corrige las señales anterógradas que envía hacia los músculos la siguiente vez que se necesite ese movimiento.
LA CÉLULA Y SUS FUNCIONES
Cada una de los 100 billones de células de un ser humano es una estructura viva que puede sobrevivir durante meses o incluso años, siempre que los líquidos de su entorno contengan los nutrientes apropiados. Las células son los elementos básicos que conforman el organismo.
- aportan la estructura de los tejidos y los órganos del cuerpo.
- ingieren los nutrientes y los convierten en energía.
- realizan funciones especializadas.
- contienen el código hereditario del organismo que controla las sustancias sintetizadas por las células y les permite realizar copias de sí mismas.
Organización de la célula
Sus dos partes más importantes son el núcleo y el citoplasma, que están separados entre sí por una membrana nuclear, mientras que el citoplasma está separado de los líquidos circundantes por una membrana celular que también se conoce como membrana plasmática.

Las diferentes sustancias que componen la célula se conocen colectivamente como protoplasma. El protoplasma está compuesto principalmente por cinco sustancias: agua, electrólitos, proteínas, lípidos e hidratos de carbono.
Agua :El principal medio líquido de la célula es el agua, que está presente en la mayoría de las células, excepto en los adipocitos, en una concentración del 70-85%. Muchos de los componentes químicos de la célula están disueltos en el agua, mientras que otros están en suspensión como micropartículas sólidas.
Iones : Algunos de los iones importantes de la célula son el potasio, el magnesio, el fosfato, el sulfato, el bicarbonato y cantidades más pequeñas de sodio, cloruro y calcio. Los iones son los productos químicos inorgánicos de las reacciones celulares y además son necesarios para el funcionamiento de algunos de los mecanismos de control celulares.
Proteínas: Después del agua, las sustancias más abundantes en la mayoría de las células son las proteínas, que normalmente constituyen entre el 10 y el 20% de la masa celular. Son de dos tipos:
1)Proteínas estructurales: están presentes en la célula principalmente en forma de filamentos largos que son polímeros de muchas moléculas proteicas individuales. Un uso importante de este tipo de filamentos intracelulares es la formación de microtúbulos que proporcionan los «citoesqueletos» de orgánulos celulares como los cilios, axones nerviosos, husos mitóticos de las células en mitosis y masas arremolinadas de túbulos filamentosos finos que mantienen unidas las partes del citoplasma y nucleoplasma en sus compartimientos respectivos.
2)Proteínas funcionales: compuesto habitualmente por combinaciones de pocas moléculas en un formato tubular-globular. A menudo son móviles dentro del líquido celular. Las enzimas entran en contacto directo con otras sustancias del líquido celular y catalizan reacciones químicas intracelulares específicas.

Lípidos : son varios tipos de sustancias que se agrupan porque tienen una propiedad común de ser solubles en disolventes grasos. Lípidos especialmente importantes son los fosfolípidos y el colesterol, que juntos suponen solo el 2% de la masa total de la célula. Su importancia radica en que, al ser principalmente insolubles en agua, se usan para formar las barreras de la membrana celular y de la membrana intracelular que separan los distintos compartimientos celulares.
Hidratos de carbono: tienen escasas funciones estructurales en la célula, salvo porque forman parte de las moléculas glucoproteicas, pero sí tienen un papel muy importante en la nutrición celular. Los hidratos de carbono siempre están presentes en forma de glucosa disuelta en el líquido extracelular circundante, de forma que es fácilmente accesible a la célula.
Estructura física de la célula
La célula contiene estructuras físicas muy organizadas que se denominan orgánulos intracelulares.
Estructuras membranosas de la célula
La mayoría de los orgánulos de la célula están cubiertos por membranas compuestas principalmente por lípidos y proteínas. Estas membranas son la membrana celular, nuclear, del retículo endoplásmico y las de la mitocondria, los lisosomas y el aparato de Golgi. Los lípidos de las membranas proporcionan una barrera que impide el movimiento de agua y sustancias hidrosolubles desde un compartimiento celular a otro, porque el agua no es soluble en lípidos.
Membrana celular
La membrana celular o membrana plasmática cubre la célula y es una estructura elástica, fina y flexible que tiene un grosor de tan solo 7,5 a 10 nm. Está formada casi totalmente por proteínas y lípidos, con una composición aproximada de un 55% de proteínas, un 25% de fosfolípidos, un 13% de colesterol, un 4% de otros lípidos y un 3% de hidratos de carbono.

La barrera lipídica de la membrana celular impide la penetración de sustancias hidrosolubles
Su estructura básica consiste en una bicapa lipídica, una película fina de doble capa de lípidos, cada una de las cuales contiene una sola molécula de grosor y rodea de forma continua toda la superficie celular. En esta película lipídica se encuentran intercaladas grandes proteínas globulares. La bicapa lipídica básica está formada por tres tipos principales de lípidos:
Fosfolípidos: son los más abundantes en la membrana celular. Un extremo de cada molécula de fosfolípido es soluble en agua, es decir, es hidrófilo (extremo fosfato), mientras que el otro es soluble solo en grasas, es decir, es hidrófobo( porción del ácido graso). La capa lipídica de la zona media de la membrana es impermeable a las sustancias hidrosolubles habituales, como iones, glucosa y urea. Por el contrario, las sustancias hidrosolubles, como oxígeno, dióxido de carbono y alcohol, pueden penetrar en esta porción de la membrana con facilidad.
Esfingolípidos: derivados del aminoalcohol esfingosina, tienen también grupos hidrófobos e hidrófilos y están presentes en pequeñas cantidades en las membranas celulares, especialmente en las células nerviosas. Los esfingolípidos complejos de las membranas celulares tienen varias funciones, como son la protección frente a factores perniciosos del entorno, la transmisión de señales y como sitios de adhesión para proteínas extracelulares.
Colesterol: Las moléculas de colesterol de la membrana son también lípidos, porque sus núcleos esteroides son muy liposolubles. Estas moléculas, en cierto sentido, están disueltas en la bicapa de la membrana. Una de sus funciones más importantes consiste en determinar el grado de permeabilidad (o impermeabilidad) de la bicapa ante los componentes hidrosolubles de los líquidos del organismo. También controla gran parte de la fluidez de la membrana.
PROTEÍNAS DE LA MEMBRANA CELULAR INTEGRALES Y PERIFÉRICAS
Estas proteínas de membrana son principalmente glucoproteínas. Existen dos tipos de proteínas de membrana celular: proteínas integrales que protruyen por toda la membrana y proteínas periféricas que se unen solo a una superficie de la membrana y que no penetran en todo su espesor. Muchas de las proteínas integrales componen canales estructurales (o poros) a través de los cuales las moléculas de agua y las sustancias hidrosolubles, especialmente los iones, pueden difundir entre los líquidos extracelular e intracelular. Otras proteínas integrales actúan como proteínas transportadoras de sustancias que, de otro modo, no podrían penetrar en la bicapa lipídica. Las proteínas integrales de la membrana pueden actuar también como receptores de los productos químicos hidrosolubles. La interacción de los receptores de la membrana celular con ligandos específicos que se unen al receptor provoca cambios conformacionales de la proteína del receptor. Las moléculas proteicas periféricas se unen con frecuencia a las proteínas integrales, de forma que las proteínas periféricas funcionan casi totalmente como enzimas o como controladores del transporte de sustancias a través de los «poros» de la membrana celular.
Hidratos de carbono de la membrana: «glucocáliz» celular
Los hidratos de carbono de la membrana se presentan casi invariablemente combinados con proteínas o lípidos en forma de glucoproteínas o glucolípidos. De hecho, la mayoría de las proteínas integrales son glucoproteínas y aproximadamente la décima parte de las moléculas lipídicas de la membrana son glucolípidos. Hay muchos otros compuestos de hidratos de carbono, que se denominan proteoglucanos y son principalmente hidratos de carbono unidos a núcleos de proteínas pequeñas, que se unen laxamente a la superficie externa de la pared celular, es decir, toda la superficie externa de la célula a menudo contiene un recubrimiento débil de hidratos de carbono que se conoce como glucocáliz.
CITOPLASMA Y SUS ORGÁNULOS
El citoplasma está lleno de partículas diminutas y grandes y orgánulos dispersos. La porción de líquido gelatinoso del citoplasma en el que se dispersan las partículas se denomina citosol y contiene principalmente proteínas, electrólitos y glucosa disueltos. En el citoplasma se encuentran dispersos glóbulos de grasa neutra, gránulos de glucógeno, ribosomas, vesículas secretoras y cinco orgánulos especialmente importantes: el retículo endoplásmico, el aparato de Golgi, las mitocondrias, los lisosomas y los peroxisomas.
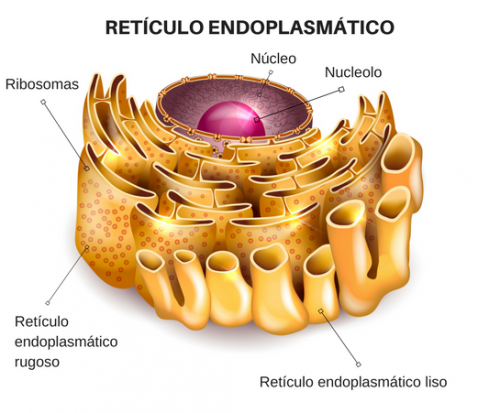
Retículo endoplásmico:
Red de estructuras vesiculares tubulares y planas del citoplasma que forman el retículo endoplásmico. Ayuda a procesar las moléculas formadas por la célula y las transporta a sus destinos específicos dentro o fuera de la célula.
Ribosomas y retículo endoplásmico rugoso:
Unidas a la superficie exterior de muchas partes del retículo endoplásmico encontramos una gran cantidad de partículas granulares diminutas que se conocen como ribosomas. Cuando estas partículas están presentes, el retículo se denomina retículo endoplásmico rugoso. Los ribosomas están formados por una mezcla de ARN y proteínas y su función consiste en sintetizar nuevas moléculas proteicas en la célula.

Retículo endoplásmico agranular:
Parte del retículo endoplásmico no tiene ribosomas, es lo que se conoce como retículo endoplásmico agranular, o liso. Este retículo agranular actúa en la síntesis de sustancias lipídicas y en otros procesos de las células que son promovidos por las enzimas intrarreticulares.
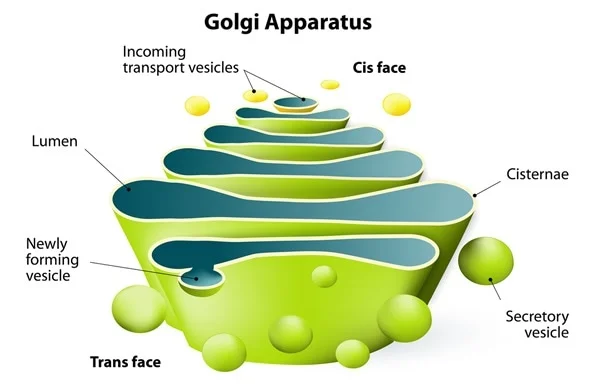
Aparato de Golgi:
Está íntimamente relacionado con el retículo endoplásmico. Tiene unas membranas similares a las del retículo endoplásmico agranular y está formado habitualmente por cuatro o más capas apiladas de vesículas cerradas, finas y planas, que se alinean cerca de uno de los lados del núcleo. Este aparato es prominente en las células secretoras, donde se localiza en el lado de la célula a partir del cual se extruirán las sustancias secretoras. Hay pequeñas «vesículas de transporte» (también denominadas vesículas del retículo endoplásmico) que continuamente salen del retículo endoplásmico y que poco después se fusionan con el aparato de Golgi. De esta forma, las sustancias atrapadas en las vesículas del RE se transportan desde el retículo endoplásmico hacia el aparato de Golgi. Las sustancias transportadas se procesan después en el aparato de Golgi para formar lisosomas, vesículas secretoras y otros componentes citoplásmicos.
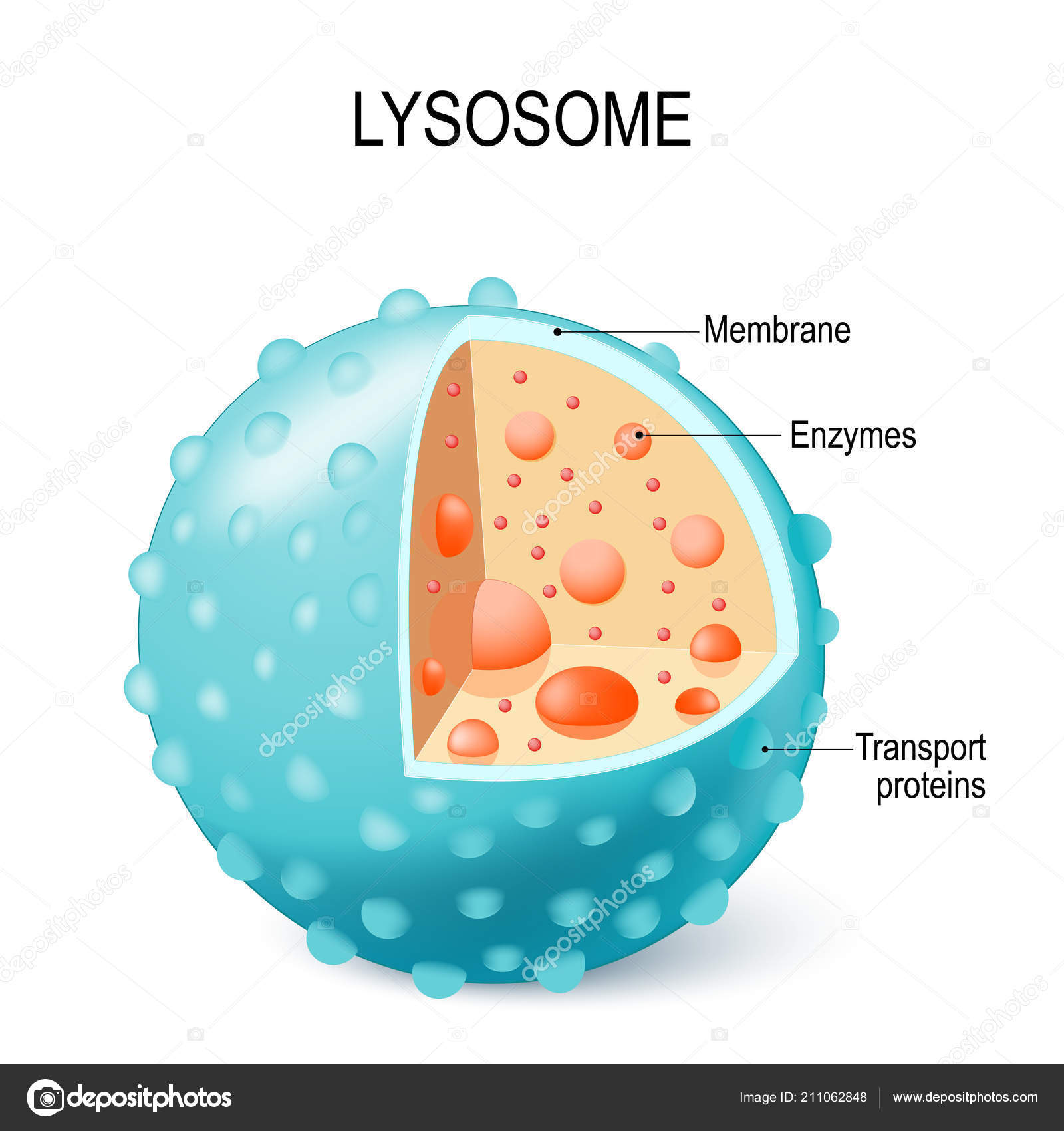
Lisosomas:
Son orgánulos vesiculares que se forman por la rotura del aparato de Golgi y después se dispersan por todo el citoplasma. Los lisosomas constituyen el aparato digestivo intracelular que permite que la célula digiera:
1) las estructuras celulares dañadas
2) las partículas de alimento que ha ingerido
3) las sustancias no deseadas, como las bacterias.
Está rodeado por una membrana bicapa lipídica típica llena con grandes cantidades de gránulos pequeños, de 5 a 8 nm de diámetro, que son agregados de proteínas que contienen hasta 40 tipos diferentes de enzimas (digestivas) de tipo hidrolasa.
Peroxisomas:
Son físicamente similares a los lisosomas; se cree que están formados por autorreplicación en lugar de proceder del aparato de Golgi. Contienen oxidasas en lugar de hidrolasas. Varias de estas oxidasas son capaces de combinar el oxígeno con los iones hidrógeno derivados de distintos productos químicos intracelulares para formar peróxido de hidrógeno (H2O2).
Vesículas secretoras:
Casi todas las sustancias secretoras se forman en el sistema retículo endoplásmico-aparato de Golgi y después se liberan desde el aparato de Golgi hacia el citoplasma en forma de vesículas de almacenamiento que se conocen como vesículas secretoras o gránulos secretores.
Mitocondrias:
Sin ellas, las células no serían capaces de extraer energía suficiente de los nutrientes y, en esencia, cesarían todas las funciones celulares. Se encuentran en todas las zonas del citoplasma de la célula, pero su número total en cada célula varía de menos de cien hasta varios miles, dependiendo de la cantidad de energía que requiere la célula. Tienen una forma y tamaño variables. La estructura básica de la mitocondria está compuesta principalmente por dos membranas de bicapa lipídica-proteínas: una membrana externa y una membrana interna. La cavidad interna de la mitocondria está llena con una matriz que contiene grandes cantidades de enzimas disueltas que son necesarias para extraer la energía de los nutrientes. La energía liberada se usa para sintetizar una sustancia de «alta energía» que se denomina trifosfato de adenosina (ATP).

Citoesqueleto celular:
estructuras filamentosas y tubulares. Es una red de proteínas fibrilares organizadas habitualmente en filamentos o túbulos que se originan como moléculas proteicas precursoras sintetizadas por los ribosomas en el citoplasma. Una de las funciones principales de los microtúbulos es actuar como citoesqueleto, proporcionando estructuras físicas rígidas para determinadas partes de las células. El citoesqueleto de la célula determina la forma celular, además participa en la división de las células, permite su movimiento y proporciona una especie de ruta que dirige el movimiento de los orgánulos en el interior de las células.
Núcleo:
Es el centro de control de la célula, envía mensajes a esta para que crezca y madure, se replique o muera. Contiene grandes cantidades de ADN, que comprende los genes, que son los que determinan las características de las proteínas celulares, como las proteínas estructurales, y también las enzimas intracelulares que controlan las actividades citoplásmicas y nucleares.
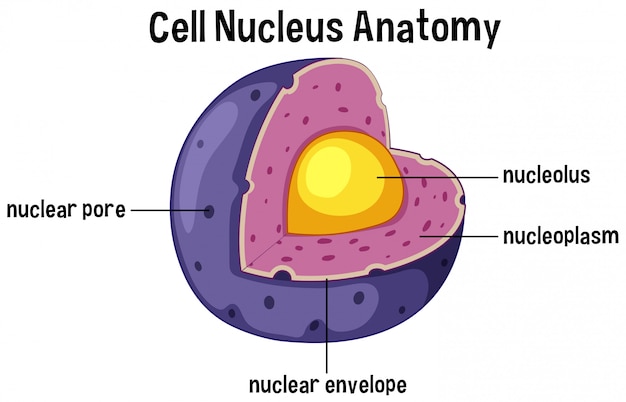
Membrana nuclear:
consiste en dos membranas bicapa separadas, una dentro de la otra. La membrana externa es una continuación del retículo endoplásmico del citoplasma celular y el espacio que queda entre las dos membranas nucleares también es una continuación con el espacio del interior del retículo endoplásmico. Varios miles de poros nucleares atraviesan la membrana nuclear.
Nucléolos y formación de ribosomas: Los núcleos de la mayoría de las células contienen una o más estructuras que se tiñen intensamente y se denominan nucléolos. Estos nucléolos no tienen una membrana limitante, sino que consisten en una acumulación simple de grandes cantidades de ARN y proteínas de los tipos encontrados en los ribosomas. La formación de los nucléolos (y de los ribosomas del citoplasma fuera del núcleo) comienza en el núcleo. Primero, los genes específicos de ADN de los cromosomas dan lugar a la síntesis de ARN, parte del cual se almacena en los nucléolos, aunque la mayoría se transporta hacia fuera, a través de los poros nucleares, hacia el citoplasma, donde se usan junto con proteínas específicas para ensamblar los ribosomas «maduros» que tienen un papel esencial en la formación de las proteínas del citoplasma.
SISTEMAS FUNCIONALES DE LA CÉLULA
Ingestión por la célula: endocitosis
Si una célula va a vivir, crecer y reproducirse, debe obtener nutrientes y otras sustancias de los líquidos circundantes. La mayoría de estas sustancias atraviesan la membrana celular por difusión y transporte activo.
*Difusión: implica el movimiento simple a través de la membrana, provocado por el movimiento aleatorio de las moléculas de la sustancia; las sustancias se desplazan a través de los poros de la membrana celular o, en el caso de las sustancias liposolubles, a través de la matriz lipídica de la membrana.
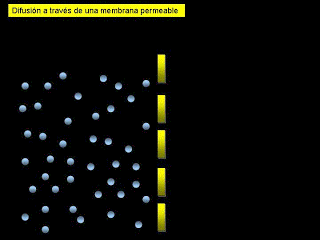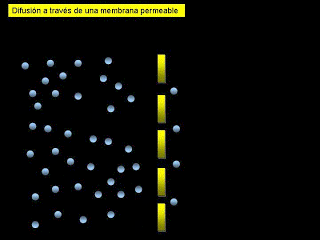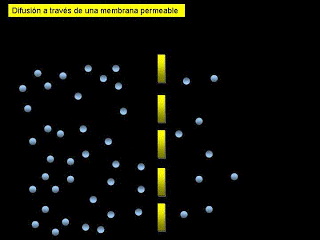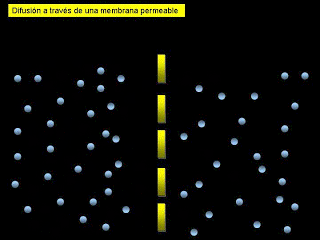
*Transporte activo: implica el transporte real de una sustancia a través de la membrana mediante una estructura física de carácter proteico que penetra en todo el espesor de la membrana. Las partículas muy grandes entran en la célula mediante una función especializada de la membrana celular que se denomina endocitosis. Las formas principales de endocitosis son:

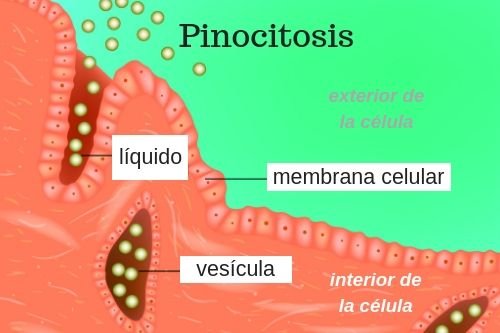
Pinocitosis: ingestión de partículas diminutas que forman vesículas de líquido extracelular y partículas dentro del citoplasma celular. se produce continuamente en las membranas celulares de la mayoría de las células, pero es especialmente rápida en algunas de ellas.
Fagocitosis: ingestión de partículas grandes, como bacterias, células enteras o porciones de tejido degenerado. Solo algunas células tienen la capacidad de realizar la fagocitosis, principalmente los macrófagos tisulares y algunos leucocitos sanguíneos.

Digestión de las sustancias extrañas introducidas por pinocitosis y fagocitosis dentro de la célula por los lisosomas
Casi inmediatamente después de que aparezca una vesícula de pinocitosis o fagocitosis dentro de una célula se unen a ella uno o más lisosomas que vacían sus hidrolasas ácidas dentro de ella. Es decir, se forma una vesícula digestiva dentro del citoplasma celular en la que las hidrolasas comienzan a hidrolizar las proteínas, los hidratos de carbono, los lípidos y otras sustancias de la vesícula. Los productos de digestión son moléculas pequeñas de aminoácidos, glucosa, fosfatos, etc., que pueden difundir a través de la membrana de las vesículas hacia el citoplasma. Lo que queda en la vesícula digestiva, que se denomina cuerpo residual, representa las sustancias indigestibles. En la mayoría de los casos, el cuerpo residual se excreta finamente a través de la membrana celular en un proceso que se denomina exocitosis, que es esencialmente lo contrario que la endocitosis.
REGRESIÓN DE LOS TEJIDOS Y AUTÓLISIS DE LAS CÉLULAS DAÑADAS
REGRESIÓN: Los tejidos del organismo a menudo regresan a un tamaño más pequeño. Ejemplos: en el útero después del embarazo, en los músculos tras períodos prolongados de inactividad y en las glándulas mamarias al final de la lactancia. Los lisosomas son responsables de gran parte de esta regresión.
ELIMINACIÓN DE LAS CÉLULAS O PORCIONES DE CÉLULAS DAÑADAS EN LOS TEJIDOS: El daño de una célula causado por el calor, el frío, un traumatismo, productos químicos o cualquier otro factor induce la rotura de los lisosomas. Las hidrolasas liberadas comienzan inmediatamente a digerir las sustancias orgánicas circundantes. Si el daño es pequeño, solo se eliminará una porción de la célula, que después se repararía. Si el daño es importante se digiere toda la célula, lo que se denomina autólisis. De esta manera, la célula se elimina por completo y se forma una célula nueva del mismo tipo, normalmente por la reproducción mitótica de una célula adyacente para ocupar el puesto de la anterior. Los lisosomas contienen sustancias bactericidas que pueden matar a las bacterias fagocitadas antes de que provoquen daños a la célula :lisozima, lisoferrina y el medio ácido. metabólicos bacterianos.
Reciclado de los orgánulos celulares: autofagia
Los lisosomas desempeñan un papel fundamental en el proceso de autofagia(comerse a sí mismo). AUTOFAGIA: proceso de limpieza según el cual los orgánulos y los grandes agregados proteicos obsoletos se degradan y se reciclan. Los orgánulos celulares deteriorados son transferidos a lisosomas por estructuras de doble membrana denominadas autofagosomas, que se forman en el citosol. Este proceso contribuye a la renovación rutinaria de los componentes citoplásmicos y es un mecanismo clave para: el desarrollo tisular, para la supervivencia celular en situaciones de escasez de nutrientes y para el mantenimiento de la homeostasis.
SÍNTESIS DE ESTRUCTURAS CELULARES EN EL RETÍCULO ENDOPLÁSMICO Y EL APARATO DE GOLGI
Estas estructuras se forman principalmente en las membranas de bicapa lipídica similares a la membrana celular y sus paredes se cargan de enzimas proteicas que catalizan la síntesis de muchas sustancias que necesita la célula. La mayor parte de la síntesis comienza en el retículo endoplásmico. Los productos formados pasan entonces al aparato de Golgi, donde también se procesan antes de ser liberados en el citoplasma.
FUNCIONES ESPECÍFICAS DEL RETÍCULO ENDOPLÁSMICO
Las proteínas se forman en el retículo endoplásmico rugoso
La porción granular del retículo endoplásmico se caracteriza por un gran número de ribosomas unidos a las superficies externas de la membrana del retículo endoplásmico. Las moléculas proteicas se sintetizan en el interior de las estructuras de los ribosomas, que extruyen parte de las moléculas proteicas sintetizadas directamente hacia el citosol, pero también extruyen muchas más moléculas a través de la pared del retículo endoplásmico hacia el interior de las vesículas y túbulos endoplásmicos, es decir, hacia la matriz endoplásmica.
Síntesis de lípidos en el retículo endoplásmico liso
El retículo endoplásmico también sintetiza lípidos, especialmente fosfolípidos y colesterol. Estos lípidos se incorporan rápidamente a la bicapa lipídica del propio retículo endoplásmico provocando que su crecimiento sea aún mayor. Para evitar que el retículo endoplásmico crezca más allá de las necesidades de la célula, las vesículas pequeñas conocidas como vesículas RE o vesículas de transporte se separan continuamente del retículo liso; la mayoría migra después rápidamente hacia el aparato de Golgi.
Otras funciones del retículo endoplásmico
1. Proporciona las enzimas que controlan la escisión del glucógeno cuando se tiene que usar el glucógeno para energía.
2. Proporciona una gran cantidad de enzimas que son capaces de detoxificar las sustancias, como los fármacos, que podrían dañar la célula. Consigue la detoxificación por: coagulación, oxidación, hidrólisis, conjugación con ácido glucurónico y de otras formas.
FUNCIONES ESPECÍFICAS DEL APARATO DE GOLGI
Funciones de síntesis del aparato de Golgi
1)procesar todavía más las sustancias que ya se han formado en el retículo endoplásmico.
2)capacidad de sintetizar ciertos hidratos de carbono que no se pueden formar en el retículo endoplásmico(ácido hialurónico y el sulfato de condroitina).
LA MITOCONDRIA EXTRAE ENERGÍA DE LOS NUTRIENTES
Las sustancias principales a partir de las cuales las células extraen energía son los alimentos, que reaccionan químicamente con el oxígeno: los hidratos de carbono, las grasas y las proteínas. En el cuerpo humano, esencialmente todos los hidratos de carbono se convierten en glucosa, lasproteínas se convierten en aminoácidos y las grasas en ácidos grasos. Dentro de la célula los alimentos reaccionan químicamente con el oxígeno, bajo la influencia de las enzimas que controlan las reacciones y canalizan la energía liberada en la dirección adecuada. casi todas estas reacciones oxidativas se producen dentro de la mitocondria y la energía que se libera se usa para formar el compuesto de alta energía ATP.
CARACTERÍSTICAS FUNCIONALES DEL ATP
El ATP es un nucleótido compuesto por: la base nitrogenada adenina, el azúcar pentosa ribosa y tres radicales fosfato. Los dos últimos radicales fosfato están conectados con el resto de la molécula mediante los denominados enlaces de fosfato de alta energía. Cuando el ATP libera su energía se separa un radical de ácido fosfórico y se forma difosfato de adenosina (ADP). La energía liberada se usa para dar energía a muchas de las demás funciones celulares, como la síntesis de sustancias y la contracción muscular. Para reconstituir el ATP celular conforme se consume, la energía derivada de los nutrientes celulares hace que el ADP y el ácido fosfórico se recombinen para formar una nueva molécula de ATP y todo el proceso se repite una y otra vez. Por este motivo, el ATP se conoce como la moneda energética de la célula porque se puede gastar y recomponer continuamente, con un ciclo metabólico de solo unos minutos.
Usos del ATP para las funciones celulares
La energía del ATP se usa para promover tres categorías principales de funciones celulares: 1) transporte de sustancias a través de múltiples membranas en la célula
2) síntesis de compuestos químicos a través de la célula
3) trabajo mecánico.
El principal uso final del ATP consiste en suministrar energía para las células especiales para realizar trabajo mecánico. Otras células realizan un trabajo mecánico de otra forma, en especial por el movimiento ciliar y amebiano.
Locomoción de las células: El tipo más evidente de movimiento que se produce en el organismo es el de los miocitos en el músculo esquelético, cardíaco y liso, que constituye casi el 50% de toda la masa del organismo.
Movimiento amebiano: es el movimiento de toda la célula en relación con su entorno, como el movimiento de los leucocitos a través de los tejidos. El movimiento amebiano comienza con la protrusión de un seudópodo desde un extremo de la célula. Este seudópodo se proyecta lejos de la célula y se asegura parcialmente en una zona nueva. Después, tira del resto de la célula hacia él.
Mecanismo de locomoción amebiana: Es consecuencia de la formación continua de una membrana celular nueva en el extremo director del seudópodo y la absorción continua de la membrana en las porciones media y posterior de la célula.
Tipos de células que muestran movimiento amebiano: Las células más frecuentes que muestran movimiento amebiano en el cuerpo humano son los leucocitos cuando salen de la sangre hacia los tejidos para formar macrófagos tisulares. Los fibroblastos se mueven hacia una zona dañada para reparar el daño e incluso las células germinales de la piel que, aunque normalmente son células totalmente sésiles, se desplazan hacia la zona de un corte para reparar el desgarro.
Cilios y movimientos ciliares:


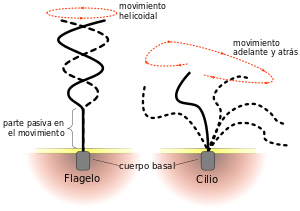
un cilio tiene el aspecto de un pelo recto o curvo con punta afilada que se proyecta 2-4 μm desde la superficie de la célula. El cilio está cubierto por una protrusión de la membrana celular y se apoya en 11 microtúbulos, 9 túbulos dobles situados en la periferia del cilio y 2 túbulos sencillos hacia el centro. El movimiento ciliar, es un movimiento a modo de látigo de los cilios que se encuentran en la superficie de las células. Este movimiento existe principalmente solo en dos lugares del cuerpo humano: en la superficie de las vías aéreas y en la superficie interna de las trompas uterinas (trompas de Falopio) del aparato reproductor. El flagelo de un espermatozoide es similar a un cilio; de hecho, tiene el mismo tipo de estructura y el mismo tipo de mecanismo contráctil. Sin embargo, este flagelo es mucho más largo y se desplaza con ondas de tipo cuasi-sinusoidal en lugar de movimientos de tipo látigo.
CONTROL GENÉTICO DE LA SÍNTESIS PROTEICA, LAS FUNCIONES DE LA CÉLULA Y LA REPRODUCCIÓN CELULAR

Los genes que están situados en el núcleo de todas las células del organismo controlan la herencia de padres a hijos y controlan la función cotidiana de todas las células del organismo.

Los genes controlan las funciones de la célula determinando qué sustancias se sintetizan dentro de esta, es decir, qué estructuras, qué enzimas y qué productos químicos participan. Cada gen (compuesto por ácido desoxirribonucleico (ADN)), controla automáticamente la formación de otro ácido nucleico, el ácido ribonucleico (ARN), que después se dispersa por toda la célula para controlar la formación de una proteína específica.
El proceso completo, desde la transcripción del código genético en el núcleo hasta la traducción del código del ARN y la formación de proteínas en el citoplasma celular, se refiere a menudo como EXPRESIÓN GÉNICA.
Dado que hay aproximadamente 30.000 genes diferentes en cada célula, es posible formar un gran número de proteínas celulares distintas. Las moléculas de ARN transcritas a partir del mismo segmento de ADN (es decir, el mismo gen) pueden ser procesadas por la célula en más de una forma, para dar origen a versiones alternativas de la proteína. El número total de diferentes proteínas producidas por los distintos tipos de células humanas se estima en al menos 100.000. Algunas de las proteínas celulares son proteínas estructurales, que, asociadas a varios lípidos e hidratos de carbono, forman las estructuras de los distintos orgánulos intracelulares. La mayoría de las proteínas son enzimas que catalizan las distintas reacciones químicas en las células.
Los genes en el núcleo celular controlan la síntesis de las proteínas
En el núcleo celular hay un gran número de genes unidos por sus extremos, formando las moléculas de doble hélice largas de ADN que tienen un peso molecular que se mide por miles de millones.

Bloques básicos de ADN
Estos compuestos químicos básicos implicados en la formación del ADN:
1) ácido fosfórico
2) azúcar desoxirribosa
3)cuatro bases nitrogenadas (dos purínicas: adenina y guanina, y dos pirimidínicas: timina y citosina).
El ácido fosfórico y la desoxirribosa forman las dos hebras helicoidales que sirven de soporte para la molécula de ADN, mientras que las bases nitrogenadas se apoyan entre las dos hebras y se conectan entre sí.


Nucleótidos
La primera etapa en la formación del ADN consiste en combinar una molécula de ácido fosfórico, una molécula de desoxirribosa y una de las cuatro bases para formar un nucleótido ácido. De esta forma se crean cuatro nucleótidos distintos, uno para cada una de las cuatro bases (ácidos desoxiadenílico, desoxitimidílico, desoxiguanílico y desoxicitidílico).
Los nucleótidos se organizan para formar dos hebras de ADN unidas laxamente entre sí
Se une un gran número de nucleótidos entre sí para formar dos hebras de ADN. A su vez, las dos hebras se unen laxamente entre sí mediante enlaces débiles como líneas discontinuas centrales.
- El esqueleto de cada hebra de ADN está compuesto por moléculas de ácido fosfórico y desoxirribosa que se van alternando.
- Las bases de purina y pirimidina se unen a los lados de las moléculas de desoxirribosa.
- Las dos hebras respectivas de ADN se mantienen unidas mediante enlaces débiles de hidrógeno (líneas discontinuas) entre las bases purínicas y pirimidínicas.
- Cada base purínica de adenina de una hebra siempre se une con una base pirimidínica de timina de la otra.
- Cada base purínica de guanina siempre se une con una base pirimidínica de citosina.
Debido a la laxitud de los enlaces de hidrógeno, las dos hebras se separan con facilidad y lo hacen muchas veces cuando realizan sus funciones en la célula. En cada vuelta completa de la hélice de la molécula de ADN hay 10 pares de nucleótidos.

Código genético
La importancia del ADN se debe a su capacidad para controlar la formación de las proteínas en la célula, que se consigue mediante un código genético. Cuando las dos hebras de la molécula de ADN se separan quedan expuestas las bases purínicas y pirimidínicas proyectándose a un lado de cada hebra de ADN. Estas bases que se proyectan son las que forman el código genético.
El código genético consta de «tripletes» sucesivos de bases, es decir, tres bases sucesivas componen una palabra del código. Los tripletes sucesivos controlan en último término la secuencia de aminoácidos en una molécula proteica que la célula debe sintetizar.

El código de ADN del núcleo celular se transfiere al código de ARN en el citoplasma celular: proceso de transcripción
Como el ADN se encuentra en el núcleo de la célula, pero la mayoría de las funciones de la célula se realizan en el citoplasma, debe haber algún mecanismo para que los genes de ADN del núcleo controlen las reacciones químicas del citoplasma. Este control se consigue mediante la intermediación de otro tipo de ácido nucleico, el ARN, cuya formación está controlada por el ADN del núcleo.
El código se transfiere al ARN en un proceso que se conoce como transcripción. A su vez, el ARN se difunde desde el núcleo a través de los poros del núcleo al compartimiento citoplásmico, donde controla la síntesis proteica.
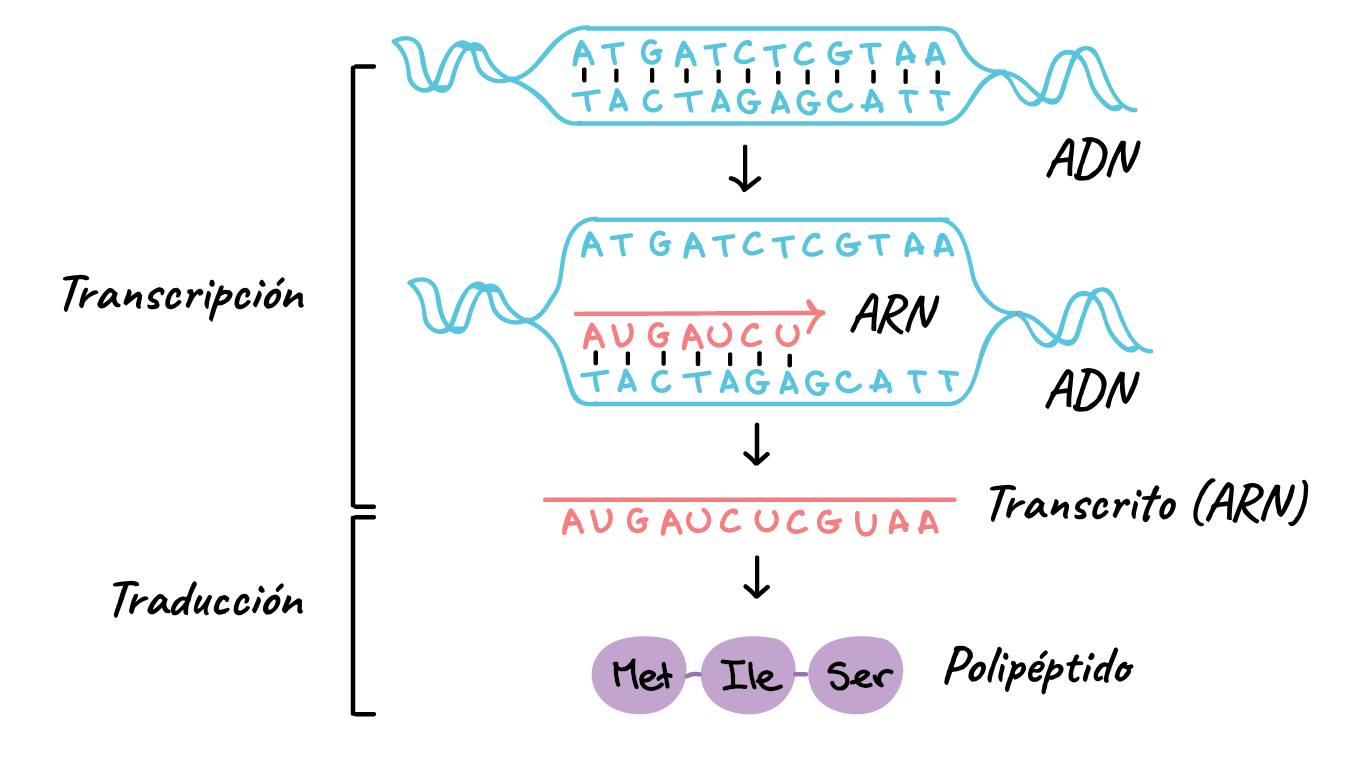
El ARN se sintetiza en el núcleo a partir de una plantilla de ADN
Durante la síntesis de ARN las dos hebras de la molécula de ADN se separan temporalmente y una de ellas se usa como plantilla para la síntesis de una molécula de ARN. Los tripletes del código del ADN provocan la formación de tripletes con un código complementario (o codones) en el ARN. A su vez, estos codones controlarán la secuencia de aminoácidos en una proteína que se va a sintetizar en el citoplasma celular.
Bloques básicos para la construcción del ARN
Los bloques básicos para la construcción del ARN son el ácido fosfórico, la azúcar ribosa, que contiene un ion hidroxilo extra unido a la estructura anular de la ribosa. Y son las mismas bases nitrogenadas, con excepción de la timina que se reemplaza por otra pirimidina, uracilo.
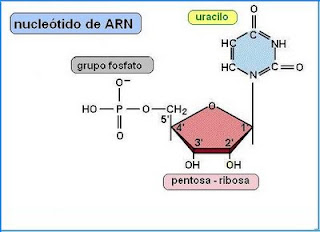
Formación de nucleótidos de ARN
Los bloques básicos de ADN forman los nucleótidos de ARN. En este caso, se usan también cuatro nucleótidos distintos para formar el ARN, nucleótidos que contienen las bases adenina, guanina, citosina y uracilo. Son las mismas bases que usa el ADN, excepto porque el uracilo del ARN reemplaza a la timina del ADN.
«Activación» de los nucleótidos de ARN
El siguiente paso de la síntesis de ARN es la «activación» de los nucleótidos de ARN por una enzima, polimerasa de ARN. Esta activación se produce añadiendo a cada nucleótido dos radicales fosfato más para formar trifosfatos. Estos dos últimos fosfatos se combinan con el nucleótido mediante enlaces de fosfato de alta energía derivados del ATP celular. El resultado de este proceso de activación es que cada uno de los nucleótidos puede disponer de grandes cantidades de energía del ATP. Esta energía se usa para favorecer las reacciones químicas que van añadiendo cada nuevo nucleótido de ARN al extremo de la cadena de ARN que se está desarrollando.
Montaje de la cadena de ARN a partir de los nucleótidos activados usando una cadena de ADN como plantilla: proceso de «transcripción»
el montaje de la molécula de ARN se realiza bajo la influencia de una enzima, la polimerasa de ARN: gran enzima proteica que tiene muchas propiedades funcionales para la formación de la molécula de ARN, como son:
1. En la cadena de ADN inmediatamente contigua al gen que se transcribirá hay una secuencia de nucleótidos denominada promotor. La polimerasa de ARN tiene una estructura complementaria apropiada que reconoce este promotor y se une a él en un paso esencial para iniciar la formación de la molécula de ARN.
2. La polimerasa de ARN, después de unirse al promotor, provoca el desenrollamiento de dos vueltas de la hélice de ADN, aproximadamente, y la separación de las porciones abiertas de las dos hebras.
3. la polimerasa se desplaza a lo largo de la hebra de ADN, desenrollando y separando temporalmente las dos hebras de ADN en cada etapa de su movimiento. A medida que se desplaza va añadiéndose en cada paso un nucleótido de ARN activado nuevo en el extremo de la cadena de ARN que se va formando, siguiendo los pasos siguientes:
a. provoca la formación de un enlace de hidrógeno entre la base del extremo de la cadena de ADN y la base de un nucleótido de ARN en el nucleoplasma.
b. la polimerasa de ARN va rompiendo dos de los tres radicales fosfato, separándolos de cada uno de estos nucleótidos de ARN y liberando grandes cantidades de energía de los enlaces de fosfato de alta energía que va rompiendo; esta energía se usa para crear el enlace covalente del fosfato que queda en el nucleótido con la ribosa en el extremo de la cadena de ARN en crecimiento.
c. Cuando la polimerasa de ARN alcanza el extremo del gen de ADN se encuentra con una secuencia nueva de nucleótidos de ADN que se conoce como secuencia terminadora de la cadena, que hace que la polimerasa y la cadena de ARN recién formada se separen de la cadena de ADN. Después, la polimerasa puede usarse una y otra vez para formar más cadenas de ARN.
d. A medida que se va formando una cadena de ARN nueva se rompen sus enlaces débiles de hidrógeno que la unen a la plantilla de ADN, porque el ADN tiene una afinidad alta para volver a unirse con su propia cadena complementaria de ADN, es decir, la cadena de ARN se separa del ADN y se libera en el nucleoplasma. Es decir, el código presente en la cadena de ADN se transmite finalmente de forma complementaria a la cadena de ARN.
Existen diferentes tipos de ARN
Los seis tipos de ARN realizan funciones distintas e independientes en la síntesis de proteínas:
1. ARN mensajero precursor (pre-ARNm): gran ARN de cadena única inmaduro que se procesa en el núcleo para formar ARN mensajero (ARNm) maduro. El pre-ARN incluye dos tipos diferentes de segmentos denominados intrones, que son eliminados por un proceso de corte y empalme (splicing), y exones, que se conservan en el ARNm final.
2. ARN nuclear pequeño (ARNnp): dirige el corte y empalme de pre-ARNm para formar ARNm.
3. ARN mensajero (ARNm): transporta el código genético al citoplasma para controlar el tipo de proteína que se forma.
4. ARN de transferencia (ARNt): transporta los aminoácidos activados a los ribosomas para usarlos en el montaje de la molécula proteica.
5. ARN ribosómico: junto con 75 proteínas distintas, forma ribosomas, las estructuras físicas y químicas en las que se montan realmente las moléculas proteicas.
6. MicroARN (ARNmi): son moléculas de ARN monocatenario de 21 a 23 nucleótidos capaces de regular la transcripción y la traducción génicas.
ARN mensajero: los codones
Las moléculas de ARN mensajero son cadenas largas y sencillas que se encuentran en suspensión en el citoplasma. Estas moléculas están compuestas por varios cientos a miles de nucleótidos de ARN en cadenas no pareadas y contienen codones que son exactamente complementarios a los tripletes del código de los genes de ADN.
Codones de ARN para los distintos aminoácidos
un codón representa la señal de «iniciar la fabricación de la molécula proteica» y tres codones representan la señal de «detener la fabricación de la molécula proteica».
ARN de transferencia: los anticodones
transfiere las moléculas de aminoácidos a las moléculas proteicas a medida que se va sintetizando la proteína. Cada tipo de ARNt se combina específicamente con 1 de los 20 aminoácidos que se van a incorporar en las proteínas.
El ARNt actúa como vehículo para transportar su tipo específico de aminoácido a los ribosomas, donde se van formando las moléculas proteicas. En los ribosomas, cada tipo específico de ARNt reconoce un codón en particular en el ARNm y libera el aminoácido apropiado en el lugar apropiado de la cadena de la molécula proteica que se está formando. Contiene solo 80 nucleótidos, es una molécula relativamente pequeña comparada con la del ARNm. En uno de los extremos de la molécula siempre hay un ácido adenílico, en cuyo grupo hidroxilo de la ribosa del ácido adenílico se une el aminoácido transportado.
Como la función del ARNt consiste en unir un aminoácido específico a una cadena de proteína en formación, es esencial que cada tipo de ARNt también sea específico de un codón en particular del ARNm. El código específico del ARNt que le permite reconocer un codón específico es, de nuevo, un triplete de bases de nucleótidos que se denomina anticodón y se sitúa aproximadamente en la zona media de la molécula del ARNt. Durante la formación de la molécula proteica las bases del anticodón se combinan laxamente mediante enlaces hidrógeno con las bases del codón del ARNm. De esta forma, los aminoácidos respectivos se alinean uno después del otro a lo largo de la cadena del ARNm, con lo que se establece la secuencia apropiada de aminoácidos de la molécula proteica que se está formando.
ARN ribosómico
El tercer tipo de ARN de la célula es el ARN ribosómico, que constituye el 60% del ribosoma. El resto está formado por proteínas, unas 75 clases distintas que son de tipo estructural, y también las enzimas necesarias para la fabricación de las moléculas proteicas.
El ribosoma es la estructura física del citoplasma en el que se sintetizan realmente las moléculas proteicas. El ribosoma actúa como una planta de fabricación en la cual se forman las moléculas proteicas.
Formación de ribosomas en el nucléolo
Los genes de ADN que se utilizan para la formación del ARN ribosómico se sitúan en cinco pares de cromosomas del núcleo. Cada uno de estos cromosomas contiene muchos duplicados de estos genes en particular, porque las funciones celulares necesitan grandes cantidades de ARN ribosómico. A medida que se forma el ARN ribosómico se recoge en el nucléolo, una estructura especializada adyacente a los cromosomas. Cuando se están sintetizando grandes cantidades de ARN ribosómico, como sucede en las células que fabrican grandes cantidades de proteínas, el nucléolo es una estructura de gran tamaño, mientras que en las células que sintetizan pocas proteínas, el nucléolo a veces ni siquiera se ve. El ARN ribosómico se procesa especialmente en el nucléolo, donde se une a las «proteínas ribosómicas» para formar productos de condensación granulares que son las subunidades primordiales de los ribosomas. Estas subunidades se liberan entonces desde el nucléolo y se transportan a través de los poros grandes de la cubierta nuclear hasta prácticamente todas las partes del citoplasma. Cuando estas subunidades entran en el citoplasma se reúnen para formar los ribosomas maduros y funcionantes. Por tanto, las proteínas se forman en el citoplasma de la célula, pero no en el núcleo celular, porque el núcleo no contiene los ribosomas maduros.
ARNmi y ARN de interferencia pequeño
Un cuarto tipo de ARN en la célula es el microARN (ARNmi), formado por cortos fragmentos de ARN monocatenario (de 21 a 23 nucleótidos) que regulan la expresión génica. Los ARNmi se codifican a partir del ADN transcrito de genes, pero no se traducen a proteínas y, por tanto, a menudo reciben el nombre de ARN no codificante. Los ARNmi son procesados por las células en moléculas que son complementarias al ARNm y que actúan para reducir la expresión génica. La generación de ARNmi supone un procesamiento especial de ARN precursores primarios más largos llamados ARNmi-pri, que son los transcriptos primarios del gen. Los ARNmi-pri se procesan a continuación en el núcleo celular por parte del complejo de microprocesador en pre-ARNmi, que son estructuras en horquilla de 70 nucleótidos. Estos pre-ARNmi son procesados después adicionalmente en el citoplasma por una enzima dicer específica que ayuda a ensamblar un complejo de silenciamiento inducido por ARN (RISC) y genera ARNmi.
Los ARNmi regulan la expresión génica por unión a la región complementaria del ARN y por la promoción de la represión de la traducción o degradación del ARNm antes de que pueda ser traducido por el ribosoma. Otro tipo de ARNmi es el ARN de interferencia pequeño (ARNsi), también denominado ARN de silenciamiento o ARN de interferencia corto. Los ARNsi son moléculas cortas de ARN bicatenario, de 20 a 25 nucleótidos de longitud, que interfieren con la expresión de genes específicos. Los ARNsi se refieren en general a ARNmi sintéticos y pueden administrarse para silenciar la expresión de genes específicos.
Formación de proteínas en los ribosomas: el proceso de «traducción»
Cuando una molécula de ARNm entra en contacto con un ribosoma, se desplaza por este a partir de un extremo predeterminado de la molécula de ARN que se especifica mediante la secuencia apropiada de las bases de ARN, el codón «iniciador de la cadena». Después, mientras el ARNm se desplaza por el ribosoma, se forma una molécula proteica, en un proceso que se conoce como traducción. Después, se marca el final de una molécula proteica cuando un codón de parada (el codón «terminador de la cadena») atraviesa el ribosoma y la molécula proteica se libera en el citoplasma.
Polirribosomas
Una molécula sencilla de ARNm puede formar moléculas proteicas en varios ribosomas al mismo tiempo, porque el extremo inicial de la cadena de ARN puede ir atravesando ribosomas sucesivos cuando abandona el primero. Las moléculas proteicas se encuentran en distintas fases de desarrollo en cada ribosoma. Como consecuencia de este proceso es frecuente encontrar agrupaciones de ribosomas, uniéndose entre 3 y 10 ribosomas a una única molécula de ARNm al mismo tiempo. Estos grupos se conocen como polirribosomas.

Muchos ribosomas se unen al retículo endoplásmico
Esta unión tiene lugar porque los extremos iniciales de muchas moléculas proteicas en formación tienen secuencias de aminoácidos que se unen inmediatamente a los locus de receptores específicos en el retículo endoplásmico; en consecuencia, estas moléculas penetran en la pared del retículo hasta la matriz del retículo endoplásmico. Este proceso otorga un aspecto granular a estas porciones del retículo en las que se están formando las proteínas y entran en la matriz.
Pasos químicos de la síntesis proteica
Los pasos de la reacción son los siguientes:
1. Cada aminoácido se activa en un proceso químico en el que el ATP se combina con el aminoácido para formar un complejo de monofosfato de adenosina con el aminoácido, con lo que se liberan dos enlaces fosfato de alta energía en el proceso.
2. El aminoácido activado, que tiene un exceso de energía, se combina entonces con su ARNt específico para formar un complejo aminoácido-ARNt y, al mismo tiempo, se libera el monofosfato de adenosina.
3. El ARNt que transporta el complejo del aminoácido entra en contacto a continuación con la molécula del ARNm en el ribosoma, donde el anticodón del ARNt se une temporalmente a su codón específico del ARNm, con lo que se alinea el aminoácido en la secuencia apropiada para formar una molécula proteica.
Después, y bajo la influencia de la enzima peptidilo transferasa (una de las proteínas del ribosoma), se forman los enlaces peptídicos entre los aminoácidos sucesivos que se van añadiendo progresivamente a la cadena proteica.
Enlace peptídico
Los aminoácidos sucesivos de la cadena de proteína se combinan entre sí según una reacción típica: al reaccionar los aminoácidos el resultado es la formación de agua y los reactivos resultantes forman una molécula única. Este proceso se conoce como enlace peptídico. A medida que se va añadiendo otro aminoácido se va formando otro enlace peptídico.
Síntesis de otras sustancias en la célula
Las miles de enzimas proteicas que se forman controlan esencialmente todas las reacciones químicas que tienen lugar en las células. Estas enzimas favorecen la síntesis de lípidos, glucógeno, purinas, pirimidinas y cientos de otras sustancias. Todas estas sustancias contribuyen a las múltiples funciones de las células.
Control de la función génica y actividad bioquímica de las células
los genes controlan tanto las funciones físicas como las químicas de las células, aunque también debe controlarse el grado de activación de los genes respectivos. Por cada gen (y hay aproximadamente 30.000 en total) hay al menos uno de estos mecanismos de retroalimentación.
Métodos de control de las actividades químicas de la célula:
1) regulación genética: se controla el grado de activación de los genes y la formación de productos génicos
2) regulación enzimática: se controlan los niveles de actividad de las enzimas ya formadas en la célula.
Regulación genética
La regulación genética, o regulación de la expresión génica, cubre todo el proceso, desde la transcripción del código genético en el núcleo hasta la formación de proteínas en el citoplasma. La regulación de la expresión génica dota a todos los organismos vivos de la capacidad para responder a los cambios en su medio. En los animales que tienen muchos tipos diferentes de células, tejidos y órganos, la regulación diferencial de la expresión génica permite también que los numerosos tipos distintos de células del cuerpo realicen cada uno funciones especializadas. La medida definitiva de la «expresión» génica es si (y en qué medida) se forman productos génicos (proteínas), ya que las proteínas se encargan de las funciones celulares especificadas por los genes. La regulación de la expresión génica puede tener lugar en cualquier punto de las rutas de transcripción, procesamiento de ARN y traducción.
El promotor controla la expresión génica
La síntesis de las proteínas celulares es un proceso complejo que se inicia con la transcripción de ADN a ARN. La transcripción de ADN está controlada por elementos reguladores encontrados en el promotor de un gen . En eucariotas, lo que incluye a todos los mamíferos, el promotor basal consta de una secuencia de siete bases (TATAAAA) denominada caja TATA, sitio de unión para la proteína de unión a TATA, y otros varios e importantes factores de transcripción que se conocen conjuntamente como complejo IID del factor de transcripción.
En la transcripción de genes en eucariotas influyen también los reforzadores, regiones de ADN que pueden unirse a factores de transcripción. Los reforzadores pueden estar situados a una gran distancia del gen sobre el que actúan o incluso en un cromosoma diferente. Asimismo pueden localizarse corriente arriba o corriente abajo del gen al que regulan. Aunque los reforzadores pueden estar ubicados a una gran distancia de su gen objeto, cuando el ADN se encuentra arrollado en el núcleo pueden hallarse relativamente cerca.
Otros mecanismos de control de la transcripción por el promotor
En las dos últimas décadas se han descubierto rápidamente variaciones del mecanismo básico de control del operón:
1. Un promotor se controla mediante el factor de transcripción situado en cualquier parte del genoma, es decir, el gen regulador provoca la formación de una proteína reguladora que, a su vez, actúe como una sustancia activadora o como un represor de transcripción.
2. En ocasiones, la misma proteína reguladora controla a la vez muchos promotores diferentes. En algunos casos, la misma proteína reguladora actúa como un activador de un promotor y como un represor de otro.
3. Algunas proteínas están controladas no en el punto de inicio de la transcripción de la cadena de ADN, sino más lejos en la misma. A veces, el control ni siquiera está en la propia cadena de ADN, sino durante el procesamiento de las moléculas de ARN en el núcleo antes de que se liberen al citoplasma; el control podría producirse también durante la formación de la proteína en el citoplasma en el curso de la traducción del ARN en los ribosomas.
4. En las células nucleadas el ADN del núcleo se envasa en unas unidades estructurales específicas, los cromosomas. Dentro de cada cromosoma el ADN se enrolla alrededor de unas proteínas pequeñas que se denominan histonas, que a su vez se mantienen unidas en un estado muy compacto mediante otras proteínas. Mientras el ADN se encuentre en este estado compactado no puede funcionar para formar ARN, aunque se están descubriendo muchos mecanismos de control que hacen que zonas seleccionadas de los cromosomas se comiencen a descompactar por partes, produciéndose la transcripción parcial del ARN. Como hay más de 30.000 genes diferentes en cada célula humana, no resulta sorprendente el gran número de formas en las que se puede controlar la actividad genética.
Los sistemas de control de los genes son especialmente importantes para controlar las concentraciones intracelulares de aminoácidos, derivados de aminoácidos y sustratos y productos intermedios del metabolismo de los hidratos de carbono, lípidos y proteínas.
El sistema genético de ADN controla la reproducción celular
La reproducción celular es otro ejemplo del papel ubicuo que tiene el sistema genético del ADN en todos los procesos vitales. Los genes y sus mecanismos reguladores determinan las características de crecimiento de las células y también si se dividen para formar nuevas células y cuándo. De esta manera, el sistema genético, tan importante, controla cada etapa del desarrollo del ser humano, desde el óvulo unicelular fertilizado hasta todo un organismo funcionante. Es decir, si hay un eje central de la vida, es el sistema genético del ADN.
Ciclo vital de la célula
El ciclo vital de una célula es el período que transcurre desde el inicio de la reproducción celular hasta el inicio de la siguiente reproducción celular. Cuando las células de los mamíferos no están inhibidas, se reproducen tan rápidamente como pueden y su ciclo vital puede ser tan solo de 10 a 30 h y termina por una serie de sucesos físicos diferenciados que se denominan mitosis y que tienen como finalidad lograr la división de la célula en dos células hijas nuevas. La mitosis en sí misma dura solo unos 30 min, por lo que más del 95% del ciclo vital de las células está representado por el intervalo entre las mitosis, o interfase, incluso en las células que se reproducen con mayor rapidez.
La reproducción celular comienza con la replicación del ADN
El primer paso consiste en la replicación (duplicación) de todo el ADN de los cromosomas. Solo después de que se haya producido esta replicación puede tener lugar la mitosis. El ADN comienza a duplicarse entre 5 y 10 h antes de la mitosis y la duplicación se completa en 4-8 h. El resultado neto es que se producen dos réplicas exactas de todo el ADN. Estas réplicas se convierten en el ADN de las dos células hijas nuevas que se formarán en la mitosis. Después de esta replicación hay otro período de 1-2 h antes de que se inicie bruscamente la mitosis. Durante este período comienzan los cambios preliminares que conducirán a la mitosis.

Fenómenos químicos y físicos de la replicación del ADN
El ADN se replica del mismo modo en que se transcribe el ARN a partir del ADN, excepto por algunas diferencias importantes:
1. Se replican las dos cadenas de ADN de cada cromosoma, y no solo una de ellas.
2. Las dos cadenas completas de la hélice de ADN se replican de extremo a extremo, y no solo algunas porciones como sucede en la transcripción del ARN. 3. Las principales enzimas que participan en la replicación del ADN componen un complejo de muchas enzimas, denominado polimerasa de ADN.
4. La formación de cada nueva cadena de ADN se produce simultáneamente en cientos de segmentos a lo largo de cada una de las dos cadenas de la hélice hasta que se replica toda la cadena. Después, la ADN ligasa une los extremos de estas subunidades.
5. Cada cadena de ADN recién formada se mantiene unida mediante un enlace débil de hidrógeno a la cadena original de ADN que se usó como plantilla, es decir, las dos hélices de ADN se enrollan unidas.
6. Como las hélices de ADN de cada cromosoma miden aproximadamente 6 cm de longitud y tienen millones de giros helicoidales, sería imposible que las dos hélices de ADN recién formadas se desenrollaran si no hubiera algún mecanismo especial.
Reparación de ADN, «corrección de lectura» y «mutaciones» del ADN
Durante la hora, más o menos, que transcurre entre la replicación del ADN y el comienzo de la mitosis hay un período de reparación muy activa y «corrección de lectura» de las cadenas de ADN; es decir, siempre que se hayan emparejado nucleótidos de ADN incorrectos con la cadena original que sirve de plantilla actúan unas enzimas especiales que cortan las zonas defectuosas y las reemplazan con los nucleótidos complementarios apropiados. Este proceso de reparación, que se consigue con las mismas polimerasas del ADN y ADN ligasas que se usan en la replicación, recibe el nombre de corrección de lectura de ADN. Debido a los procesos de reparación y corrección de lectura, el proceso de transcripción pocas veces comete errores pero, cuando lo hace, el error se denomina mutación. La mutación provoca la formación de alguna proteína anormal en la célula en lugar de la proteína necesaria, lo que conduce a funciones celulares anormales y, en ocasiones, incluso a la muerte celular. Cromosomas y su replicación

Las hélices de ADN del núcleo se enrollan en cromosomas. La célula humana contiene 46 cromosomas dispuestos en 23 pares. Los genes de los dos cromosomas de cada par son idénticos o casi idénticos entre sí en su mayor parte, por lo que se dice que en los pares también puede haber genes diferentes, aunque no sea siempre así. Además del ADN en el cromosoma hay una gran cantidad de proteínas, principalmente muchas moléculas pequeñas de histonas cargadas positivamente.
Estas histonas se organizan en un gran número de pequeños núcleos, a modo de carretes, alrededor de los cuales se enrollan secuencialmente pequeños segmentos de cada hélice de ADN. Los núcleos de histonas tienen un papel importante en la regulación de la actividad del ADN porque este no puede funcionar como plantilla para la formación de ARN ni para la replicación de ADN nuevo mientras esté fuertemente enrollado. La replicación de todos los cromosomas se produce en los minutos siguientes a la finalización de la replicación de las hélices de ADN y las hélices nuevas recogen las moléculas proteicas nuevas a medida que las van necesitando. Los dos cromosomas recién formados se mantienen unidos entre sí en un punto que se denomina centrómero, situado cerca del centro. Estos cromosomas duplicados, pero aún unidos entre sí, se conocen como cromátidas.
Mitosis celular

El proceso real por el que la célula se divide en dos células nuevas es la mitosis. Una vez que cada cromosoma se ha replicado para formar las dos cromátidas, en muchas células la mitosis se produce automáticamente en 1 o 2 h.
Aparato mitótico: función de los centríolos
Uno de los primeros pasos de la mitosis tiene lugar en el citoplasma, al final de la interfase, en torno o en unas pequeñas estructuras denominadas centríolos. Dos pares de centríolos se mantienen estrechamente unidos cerca de un polo del núcleo (estos centríolos, ADN y los cromosomas, se han replicado durante la interfase, después de la replicación del ADN). Cada centríolo es un pequeño organismo cilíndrico en torno a 0,4 μm de largo y 0,15 μm de diámetro y está formado principalmente por nueve estructuras tubulares paralelas dispuestas en forma de cilindro. Los dos centríolos de cada par se disponen en ángulos rectos entre sí y cada par de centríolos, junto con el material pericentriolar unido a él, compone el centrosoma. Poco antes de que tenga lugar la mitosis, los dos pares de centríolos comienzan a separarse uno de otro. Este movimiento se debe a la polimerización de las proteínas de los microtúbulos que crecen entre los pares respectivos de centríolos y los separan. Al mismo tiempo, crecen radialmente otros microtúbulos que alejan los pares de centríolos, formando una estrella a modo de soporte, que se conoce como áster, en cada extremo de la célula. El complejo de microtúbulos que se extiende entre los dos nuevos pares de centríolos es el huso, y todo el conjunto de microtúbulos más los dos pares de centríolos se denomina aparato mitótico.
Profase: La primera etapa de la mitosis. Mientras se forma el haz, los cromosomas del núcleo (que en la interfase corresponden a hebras laxamente enrolladas) se condensan en cromosomas bien definidos.
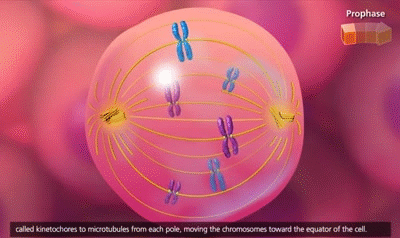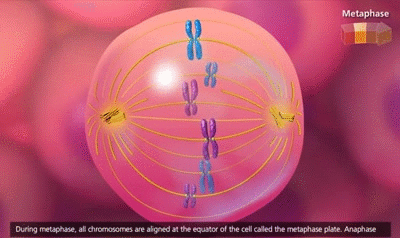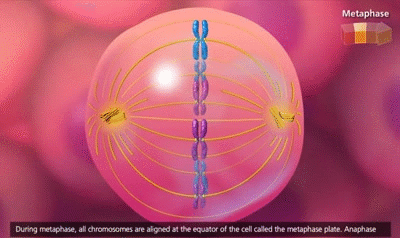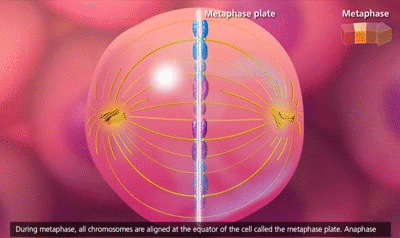
Prometafase: Las puntas de los microtúbulos en crecimiento del áster se fragmentan en la cubierta nuclear. Al mismo tiempo, los múltiples microtúbulos del áster se unen a las cromátidas en los centrómeros, donde las cromátidas pareadas aún están unidas entre sí; a continuación, los túbulos tiran de una cromátida de cada par, alejando cada una hacia el polo celular correspondiente.

Metafase: las dos ásteres del aparato mitótico se separan. Este empuje parece suceder porque las puntas de los microtúbulos de ambos, donde se imbrican entre sí para formar el huso mitótico, realmente se empujan mutuamente. Unas moléculas proteicas contráctiles diminutas, o «motores moleculares», que tal vez estén formadas por la proteína muscular actina se extienden entre las puntas respectivas para que deslicen activamente las puntas en dirección opuesta una de otra. Simultáneamente, los microtúbulos insertados en las cromátidas tiran fuertemente de ellas hasta el centro de la célula, alineándolas para formar el plano ecuatorial del huso mitótico.

Anafase: Las dos cromátidas de cada cromosoma son separadas en el centrómero. Se separan los 46 pares de cromátidas y se forman dos juegos independientes de 46 cromosomas hijos. Cada uno de ellos es empujado hacia cada uno de las ásteres de la mitosis, a medida que los dos polos respectivos de la célula en división se van separando entre sí.

Telofase: Los dos juegos de cromosomas hijos se separan completamente. A continuación, el aparato mitótico se disuelve y se desarrolla una nueva membrana nuclear que rodea cada grupo de cromosomas. Esta membrana se forma a partir de porciones del retículo endoplásmico que ya están presentes en el citoplasma. Poco después, la célula se divide en dos, en la zona media entre los dos núcleos. Este pinzamiento se produce como consecuencia de la formación de un anillo contráctil de microfilamentos compuestos por actina y, probablemente, miosina en la unión de las células nuevas que se están desarrollando, anillo que las termina separando.
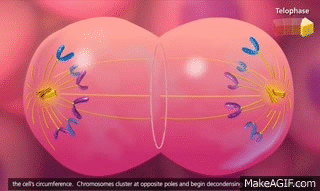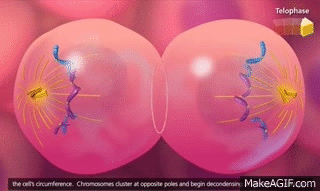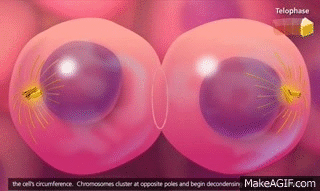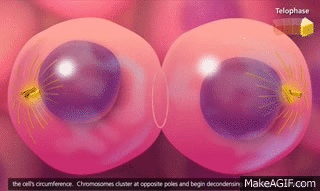
Control del crecimiento y la reproducción celular
Algunas células crecen y se reproducen continuamente, como las células que dan lugar a los elementos formes sanguíneos en la médula ósea, las capas germinales de la piel y el epitelio intestinal, mientras que muchas otras células, como los miocitos del músculo liso, pueden no reproducirse durante muchos años. Algunas células, como las neuronas y la mayoría de los miocitos del músculo estriado, no se reproducen durante toda la vida de una persona excepto durante el período de vida fetal. En algunos tejidos la falta de algunos tipos de células hace que crezcan y se reproduzcan con rapidez hasta que vuelva a haber un número apropiado de estas células. Los mecanismos que mantienen el número apropiado de los distintos tipos de células en el organismo no se conocen con detalle.
Formas de controlar el crecimiento:
1)el crecimiento se controla a menudo mediante factores de crecimiento que proceden de otras partes del organismo. Algunos de estos factores de crecimiento circulan en sangre, pero otros se originan en los tejidos adyacentes.
2)la mayoría de las células normales dejan de crecer cuando han salido de este espacio para su crecimiento, lo que sucede cuando las células crecen en un cultivo tisular; el crecimiento continúa hasta que las células entran en contacto con un objeto sólido y después se detiene.
3)las células que crecen en un cultivo celular dejan de hacerlo cuando se ha podido recoger cantidades diminutas de sus propias secreciones en el medio de cultivo. Además, este mecanismo sería un medio de controlar el crecimiento mediante la retroalimentación negativa.
Los telómeros evitan la degradación de los cromosomas
Un telómero es una región de secuencias de nucleótidos repetitivas situadas en cada extremo de una cromátida. Actúan como cubiertas protectoras que evitan que el cromosoma se deteriore durante la división celular. En el curso de la división celular, una pieza corta de ARN «cebador» se une a la cadena de ADN para iniciar la replicación. Sin embargo, dado que el cebador no se une en el extremo de la cadena de ADN, la copia está ausente en una pequeña sección del ADN. Con cada división celular, el ADN copiado pierde nucleótidos adicionales de la región del telómero. Las secuencias de nucleótidos proporcionadas por los telómeros evitan, de esta forma, la degradación de los genes cerca de los extremos de los cromosomas. Sin telómeros, los genomas perderían información progresivamente y quedarían truncados después de cada división celular. los telómeros pueden considerarse elementos cromosómicos desechables que ayudan a mantener la estabilidad de los genes, pero se consumen de forma gradual durante divisiones celulares repetidas.
Cada vez que una célula se divide, una persona media pierde de 30 a 200 pares de bases en los extremos de los telómeros de esa célula. Finalmente, cuando los telómeros se acortan hasta una longitud crítica, los cromosomas se vuelven inestables y las células mueren. Según se cree, este proceso de acortamiento de los telómeros es un motivo importante de algunos cambios fisiológicos asociados con el envejecimiento. La erosión de los telómeros puede producirse también como consecuencia de enfermedades, especialmente las asociadas con el estrés oxidativo y la inflamación.
Regulación del tamaño de la célula
El tamaño de la célula está determinado casi en su totalidad por la cantidad de ADN funcionante que hay en el núcleo. Si el ADN no se replica, la célula crece hasta un tamaño determinado y después lo mantiene, mientras que también es posible que, usando el quimioterápico colchicina, se impida la formación del huso mitótico y, por tanto, la mitosis, aunque continúe la replicación del ADN. En este caso, el núcleo contiene una cantidad de ADN bastante mayor de lo que normalmente contendría y la célula crece hasta un tamaño proporcionalmente mayor. Se supone que este crecimiento celular es consecuencia del aumento de la producción de ARN y de las proteínas celulares, lo que, a su vez, hace que el tamaño de la célula aumente más.
Diferenciación celular
se refiere a los cambios de las propiedades físicas y funcionales de las células a medida que proliferan en el embrión para formar las distintas estructuras y órganos corporales. Las células humanas maduras producen un máximo de 8.000 a 10.000 proteínas y no las 30.000 o más que se produciría si todos los genes estuvieran activos. Los estudios embriológicos demuestran que algunas células del embrión controlan la diferenciación de las células adyacentes. Por ejemplo, el mesodermo de la notocorda primordial se conoce como el organizador primario del embrión porque forma un foco alrededor del cual se desarrolla el resto del embrión. Se diferencia en un eje mesodérmico que contiene somitas de distribución segmentaria y, como consecuencia de la inducción de los tejidos circundantes, da paso a la formación de esencialmente todos los órganos del organismo.

Apoptosis: muerte celular programada
Los 100 billones de células del organismo forman parte de una comunidad muy bien organizada en la que el número total de células está regulado no solo por el control de la velocidad de división celular, sino también por el control de la velocidad de la muerte celular. Cuando las células ya no se necesitan, o cuando se convierten en una amenaza para el organismo, sufren una muerte celular programada suicida, o apoptosis: proceso que implica una cascada proteolítica específica que hace que la célula se encoja y condense para desmontar su citoesqueleto y alterar su superficie de tal forma que una célula fagocítica cercana, como un macrófago, se pueda unir a la membrana celular y digerir la célula. La apoptosis es una muerte celular metódica que da lugar al desmontaje y fagocitosis de la célula antes de que se produzca ninguna fuga de su contenido, por lo que las células vecinas se mantienen sanas.

Necrosis celular: proceso en que las células que mueren como consecuencia de una lesión aguda se hinchan y estallan debido a la pérdida de la integridad de la membrana celular. Las células necróticas vierten su contenido haciendo que la inflamación y la lesión se extiendan a las células vecinas.

Cáncer

El cáncer se debe en la mayoría de los casos a la mutación o a alguna otra activación anormal de los genes celulares que controlan el crecimiento y la mitosis celular. Los protooncogenes son genes normales que codifican diversas proteínas responsables del control de la adhesión celular, el crecimiento y la visión. Si mutan o se activan de forma excesiva, los protooncogenes pueden convertirse en oncogenes con funcionamiento anómalo capaces de provocar cáncer. En los cánceres humanos se han descubierto hasta 100 tipos distintos de oncogenes. En todas las células también hay antioncogenes, denominados también genes supresores de tumores, que suprimen la activación de los oncogenes específicos, es decir, la pérdida o inactivación de los antioncogenes permite la activación de los oncogenes que conduce al cáncer.
La mayoría de las células mutadas tiene una capacidad de supervivencia menor que las células normales y, simplemente, mueren. Solo algunas de las células mutadas que sobreviven son cancerosas, porque incluso la mayoría de las células mutadas tiene controles de retroalimentación normales que impiden su crecimiento excesivo. Estas proteínas activan el sistema inmunitario del organismo, lo que hace que generen anticuerpos o linfocitos sensibilizados que reaccionan contra las células cancerosas y las destruyen. Normalmente se necesita la presencia simultánea de varios oncogenes activados para provocar el cáncer.
Causas de alteraciones genética: la probabilidad de mutaciones aumenta enormemente cuando una persona se expone a determinados factores químicos, físicos o biológicos, como son los siguientes:
1. radiación ionizante, como los rayos X, los rayos g y la radiación de partículas procedentes de sustancias radiactivas, e incluso la luz ultravioleta, predispone al cáncer.
2. Algunas sustancias químicas también tienen una mayor propensión a provocar mutaciones. Las sustancias químicas que provocan la mutación se denominan carcinógenos.
3. Los irritantes físicos también provocan cáncer, como sucede durante la abrasión continuada del revestimiento del aparato digestivo por algunos alimentos. El daño de los tejidos conduce a una sustitución mitótica rápida de las células.
4. En muchas familias hay una importante tendencia hereditaria al cáncer.
5. Algunos virus provocan ciertas clases de cáncer, como la leucemia. Este fenómeno se consigue normalmente por una de dos vías: en el caso de los virus de ADN, la cadena de ADN de un virus se inserta a sí misma en uno de los cromosomas y, de esta forma, se provoca una mutación que conduce al cáncer; en el caso de los virus ARN, algunos transportan una enzima denominada transcriptasa inversa que provoca la transcripción del ADN desde el ARN. Este ADN transcrito se inserta en el genoma de la célula animal, con lo que se produce el cáncer.
Características invasivas de la célula cancerosa
1. La célula cancerosa no respeta los límites habituales del crecimiento celular, ya que no requieren los mismos factores de crecimiento que son necesarios para el crecimiento de las células normales.
2. Las células cancerosas son bastante menos adhesivas entre sí que las células normales, por lo que tienden a dispersarse por los tejidos, entrar al torrente sanguíneo y transportarse por el organismo, donde forman nidos para crecimientos cancerosos nuevos.
3. Algunos cánceres también producen factores angiogénicos que provocan el crecimiento de muchos vasos sanguíneos nuevos dentro del cáncer, por lo que aportan los nutrientes necesarios para el crecimiento celular.
¿Por qué matan las células cancerosas?
El tejido canceroso compite con los tejidos normales para lograr los nutrientes. Como las células cancerosas continúan proliferando indefinidamente, su número se multiplica día a día y pronto demandan prácticamente toda la nutrición disponible para el organismo o para una parte esencial de este. En consecuencia, los tejidos normales sufren una muerte nutritiva gradual.
